© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Рукопись деп. в ВИНИТИ 08.12.1987, № 8593-В87
, 63 с.КОЛИЧЕСТВЕННАЯ ОЦЕНКА ДОМИНИРОВАНИЯ В ЭКОЛОГИЧЕСКИХ СООБЩЕСТВАХ
Баканов А.И.
Институт биологии внутренних вод АН СССР, пос. Борок.
I. ВВЕДЕНИЕ. ТЕРМИНОЛОГИЯ
Экологии как науке немногим более ста лет. Начав с чисто описательных работ, в настоящее время она превратилась в одну из самых математизированных ветвей биологии (109, 153). Но возможности описательного подхода еще не исчерпаны, и современные исследования, наряду с экспериментальными и теоретическими аспектами, включают и этап количественного описания изучаемых объектов. Современная экология представляет собой учение об экосистемах, раскрывающее закономерности их состава, структуры, функционирования и эволюции (109). Для раскрытия экосистемных закономерностей необходимо иметь достаточно полное, точное, единообразно понимаемое и формализованное описание всех важнейших параметров экосистемы. Настоящая работа посвящена описанию одного из аспектов структуры экологических сообществ.
В философско-методологической литературе содержится много определений понятия “структура” различной степени абстрактности, но общепринятого определения нет. В качестве прикладного, рабочего определения примем, что структура - совокупность характеристик неоднородности изучаемого объекта. Эти характеристики могут относится к трем аспектам структуры: соотношению отдельных составных частей, взаимной связи между частями, изменению частей. В результате разностороннего изучения исследователи выделяют 10 - 20 и более типов структур (54, 91), в частности можно рассматривать таксономическую, возрастную, половую, размерную, трофическую, временную, пространственную, лимитационную, информационную, генетическую, этологическую, социальную, корреляционную и другие структуры. Разумеется, это деление достаточно условно, отдельные типы структур могут рассматриваться как часть других типов, они могут перекрываться, быть взаимосвязанными. Рассматриваемая здесь структура доминирования может в определенном смысле считаться частью таксономической структуры. Доминирование можно изучать с разных позиций, например, исследовать, почему в данном сообществе доминирует тот или иной вид, какое значение имеет характер доминирования для функционирования сообщества, как связана структура доминирования с другими характеристиками экосистемы и т. д., данная же работа посвящена методам количественной оценки доминирования, без чего невозможно его дальнейшее изучение, тем более, что в мировой литературе нет обзорных работ, специально посвященных этому вопросу.
Изучение структур сообществ имеет ряд особенностей по сравнению с изучением популяций. При исследовании популяций основным объектом является индивидуум, обычно достаточно четко отграниченный от других, размеры и признаки его поддаются измерению без существенных затруднений. Изучение же сообществ базируется на произвольно разграниченных учетных площадках, выявление сообществ представляет серьезные трудности, так как резкие границы в природе практически отсутствуют. Характеристики сообществ имеют, как правило, стохастический характер. При классификации сообществ приходится прямо или косвенно учитывать признаки не одного, а трех уровней организации живой материи: организменный, популяционный, ценотический, а также большое количество признаков среды. Закономерности и зависимости внутри сообщества обычно нелинейны. Поэтому здесь недостаточно знания биометрии, столь хорошо зарекомендовавшей себя при изучении популяций, ставится вопрос о выделении специальной отрасли науки - биоценометрии (96, 111). Кроме того, отношение доминирования нетранзитивно. Транзитивность - свойство отношений, состоящее в том, что если первый член отношения сравним со вторым, а второй с третьим. Например, если А = Б и В = С, то и А = С. Нетранзитивные же отношения не подвергались общему математическому рассмотрению, в этом направлении делаются только первые шаги. Нетранзитивность может порождаться очень большим разнообразием биологических объектов, причем даже если каждый признак сам по себе транзитивен, то их объединение может быть нетранзитивным (95). Все это затрудняет разработку количественных характеристик соответствующих структур.
В данной работе рассматривается только экологический аспект понятие “доминирование”, не касаясь того, как его трактуют в генетике ((доминантный ген), в психологии (принцип доминанты) и в этологии. В экологии это понятие иногда также употребляют в необщепринятом смысле. Например, доминантная клика - клика, не входящая в какую-либо другую клику. Клика - это группа видов внутри экосистемы, в которой каждая пара видов имеет хотя бы один общий пищевой ресурс (165). Из экологических дисциплин наибольшее внимание изучению доминирования уделяется в фитоценологии (геоботанике), например, в растительном покрове СССР выделено около 1500 доминантов, дающих основную массу растительной продукции, используемой человеком, это нашло отражение в трех томной сводке “Доминанты растительного покрова Советского Союза” (15). В фитоценологии выделяется специальный раздел - синморфология, изучающий морфологическое строение фитоценозов (70), поэтому многие принципы и показатели данной работы взяты из работ фитоценологов.
Термин “доминирование” обычно определяется через свои синонимы, толкуемые разными авторами неоднозначно: господство, значимость, лидирование, превалирование, важность, существенность, удельный вес, преобладание и т. п. Доминирующие виды называются также руководящими, основными, ключевыми, определяющими, детерминантами, предоминантами, эдификаторами, превалидами... Иногда определения представляют собой тавтологию, например, “Господство - преобладание более жизнеспособных особей над другими особями того же или другого вида” (17, стр. 55). Как неоднозначно понимается доминирование ведущими экологами видно из приводимых ниже цитат.
“Не все организмы сообщества играют одинаково важную роль в определении его природы и функций. Из сотен или тысяч видов, входящих в состав какого-либо сообщества, обычно лишь относительно немногие виды или группы оказывают на него определенное воздействие, обусловленное их численностью, размерами, продукцией или другими параметрами... Сообщества, во всяком случае основные, состоят из продуцентов, макроконсументов и микроконсументов. В пределах этих групп отдельные виды или группы видов, которые принимают значительное участие в регуляции энергетического обмена и оказывают существенное влияние на среду обитания других видов, известны как экологические доминанты... В общем доминанты - это те виды, которые на своем трофическом уровне обладают наибольшей продуктивностью... Численность, биомасса и некоторые другие показатели применяются тогда, когда сравниваемые популяции имеют примерно одинаковые соотношения размера и метаболизма. Данные по численности приводят к преувеличению значения мелких организмов, а данные по биомассе - к преувеличению роли крупных организмов. Поток энергии - более подходящий показатель” (83, стр. 109, 185, 186).
“ Доминирование означает преобладание или относительное обилие, т.е. отношение числа особей данного вида к общему числу особей всех видов, выраженное в процентах. Совершенно очевидно, что доминирование имеет биологическое значение лишь в применении к растениям, большинство которых имеют одинаковые размеры; иногда пользуются понятием ”доминирование проективного покрытия” выражаемого не числом особей, а относительной площадью поверхности, занятой видом” (62, стр. 212).
“Это понятие нельзя выразить количественно. Оно означает влияние, оказываемое каким-либо видом в любом сообществе. Один какой-то вид может быть немногочисленным, но может оказывать при этом более сильное воздействие на сообщество, чем, скажем, другой более обильно представленный вид, но меньшего размера или не очень активный. Несколько крупных жвачных окажут на луг больше влияния, чем многочисленные насекомые-фитофаги. Их-то и называют доминирующими видами. Доминирование часто выявляют в пределах какой-либо определенной систематической группы, а не всего животного мира. Следует отметить, наконец, что растительность, благодаря своему обилию является вообще группой, доминирующей над животными, за исключением некоторых биоценозов” (26, стр. 269).
“Виды, представленные в биоценозе наибольшим числом особей и биомассой, называются доминантными, лидирующими или видами-эдификаторами... Оценка значения вида в биоценозе зависит от выбора показателя количественной представленности... В последнее время все чаще степень доминантности вида оценивают по той роли, какую он играет в трансформации вещества или энергии” (47, стр. 290).
“ Для характеристики биоценотической роли видов различных организмов обычно с успехом пользуются показателями их обилия. Наиболее адекватна оценка роли в биоценозе через показатели обилия для зеленых растений, для которых она достаточно полно описывается их продукцией... Хуже характеризуют их значение в биоценозах данные о биомассе, числе особей, относительном покрытии, которые, однако, также часто используются экологами растений, так как получить их легче. Роль животных как консументов разных порядков значительно многообразнее. Определяемая в конечном счете степенью их воздействия на продуцентов, она в общем находится далеко не в прямой связи с их “продукцией”, биомассой или числом особей. Известно, что воздействие даже немногочисленных животных на определенные особенно чувствительные (критические) стадии развития растений или других животных может привести к значительным биоценотическим сдвигам. Однако изучение особенностей влияния вида животного на биоценоз возможно только в специальных и достаточно тонких исследованиях. Поэтому обычно в полевой зоологии наиболее важной биоценотической характеристикой вида считается его обилие. В общем случае термин “доминирование” нельзя признать удачным, так как доминирование имеет биологический смысл лишь при условии взаимодействия особей разных видов.... Поэтому лучше и точнее, подразумевая соотношение обилия видов в сообществе, в выборке из него и тем более в некоторой коллекции животных, вместо термина “доминирование” употреблять “численное преобладание”, а вместо “доминант” - “наиболее обильный вид” (86, стр. 22-23).
“Доминант” - вид, количественно преобладающий в данном сообществе, как правило в сравнении с близкими формами или, во всяком случае, входящими в один уровень экологической пирамиды... Доминантность (доминирование) - способность вида занимать в сообществе главенствующее положение и оказывать преобладающее влияние на ход биоценотических процессов” (92, стр. 42).
“Роль и значение вида в биосфере определяется его местом и удельным весом в биологическом круговороте веществ... Между основной пищей и ее потребителями существуют постоянные пищевые связи. С сезонной и случайной пищей связи временны и непостоянны. Первая группа составляет ядро биоценоза и называется эдификаторами или доминантами” (77, стр. 299, 529).
“Оценка важности вида зависит от цели исследования и может производится разными способами. Подобно тому, как преподаватель задает студентам несколько вопросов и по ответам оценивает их знания вообще по предмету, так исследователь может определить несколько простых параметров системы и по ним судить о состоянии ее в целом. Такие свойства, как биомасса, годовая продукция, покрытие, продукция подстилки... могут быть хорошими показателями важности. Некоторые легко измеряемые величины (плотность, частота, высота...) не прямо связаны с важностью, но, комбинируя их, можно получить удовлетворительную оценку важности” (133, стр. 794).
“...едва ли правомочен абстрактный термин” значимость организмов” в сообществе... Нам представляется, что нет оснований говорить о “роли”, “значимости” и т. д. без четкого представления тех аспектов, к которым относятся исследуемые закономерности. Необходимо различать закономерности морфологии и трофодинамических отношений - показатели “удельного веса” в совокупности морфологических частей и роль в трофодинамических процессах, степень взаимодействия и коррелятивные соотношения. Энергетические показатели, безусловно, не могут быть эквивалентом всех форм биоценотических отношений” (114, стр. 1080).
“Для высшей категории обилия обычно применяют термин “доминант”. Он довольно неудачен, но, по-видимому, слишком прижился, что бы его можно было заменить. На практике он обычно не означает ничего большего, чем ”высшая степень численности или покрытия в исследуемой растительной группировке”. Многие геоботаники... склонны смешивать такого рода использование термина “доминант” с концепцией доминантности как степени воздействия, оказываемого данным видом на другие виды в сообществе (путем различного рода конкурентных или стимулирующих влияний). Очевидно, что некоторая степень такого контролирующего влияния часто приписывается видам, числящим в списке “доминантности”, без каких либо иных доказательств, кроме оценки высоким баллом количественного участия. Вид может быть в сообществе доминантом в обоих значениях этого термина, но это вовсе не обязательно. Более того, само существование доминирования во втором значении не всегда признается. Безусловно, замена термина “доминирование” в значении количественного участия каким-либо другим, если бы это удалось произвести, способствовала бы достижению большей ясности. “Доминант” сообщества определялся бы тогда как вид, оказывающий наибольшее влияние на другие виды сообщества и испытывающий наименьшее влияние с их стороны. В геоботанической литературе встречается иногда слово “доминант” в третьем значении, когда оно означает отдельные деревья в древесном ярусе, крона которых более, чем наполовину находится на полном солнечном свету” (25, стр. 21-22).
“Доминанты - преобладающие или доминирующие в главных слоях биоценозов виды растений (виды, преобладающие во второстепенных слоях, называются субдоминантами)... Доминантность - сложившаяся в процессе биотоценогеноза способность некоторых видов занимать в биоценозе главенствующее положение и оказывать преобладающее влияние на образование в нем биоценотической среды. Доминантность вида является функцией степени его участия в сообществе и устойчивости этого участия”(17, стр. 62-63).
”Виды входящие в состав биоценоза, очень сильно различаются по свей значимости. Значение отдельных видов должно определятся тем, какую роль они играют в функционировании экосистемы или в продукционном процессе, но при исследовании биогидроценозов установить истинную функциональную роль видов нелегко, и об их значении можно в первом приближении судить по их обилию, т.е. численности и биомассе... Но одна биомасса, как и одна численность тоже не может достаточно полно охарактеризовать роль вида в биоценозе, так как в число доминантов или субдоминантов могут попасть крупные виды, найденные в большом количестве только в одной или двух точках биотопа. Поэтому необходимо привлечь еще “встречаемость” вида” (72, стр. 167, 168).
Процитированные авторы отражают почти весь спектр представлений о доминировании, встречающихся в экологической литературе. Доминирующие виды в большей степени приспособлены к условиям биотопа, причем эта адаптация - результат длительного исторического развития, поэтому некоторые авторы считают, что доминирующими можно называть только такие виды, которые сохраняют это свойство в течение достаточно продолжительного периода времени (101, 112).
При выделении доминирующих видов опираются на разные показатели: численность, биомассу (8, 47, 62, 70, 92, 112, 121), продукцию (47, 72, 77, 83, 105, 163), проективное покрытие (25, 58, 62, 70, 105), встречаемость, часто учитывается несколько показателей. Разумеется, вид, доминирующий по
одному показателю, может не быть таковым по другому. В.Д. Федоров выделил два аспекта этого термина - структурный и функциональный (106). В первом случае к доминирующим относят виды, имеющие наибольшую биомассу, численность, продукцию... т.е. которых по какому-то показателю “больше”, чем прочих видов сообщества. При втором подходе к доминирующим относят виды, наиболее сильно влияющие на другие виды сообщества, при этом сила влияния вида может быть не пропорциональна его биомассе или даже продукции. Например, часто для растений решающее значение имеет не сколько у них отчуждается фитомассы, а что отчуждается, поскольку растения особенно чувствительны к уничтожению их “точек роста” (49).Первый подход более привлекателен тогда, когда оценивается роль вида с точки зрения возможности использования его человеком. Например, доминирующий в уловах вид рыб - вид, которого выловлено наибольшее количество центнеров, доминирующие в очистном сооружении беспозвоночные - те, которые вносят наибольший вклад в очистку сточных вод. Второй подход характеризует вид с точки зрения интересов сообщества, поэтому в зависимости от цели исследования оба подхода имеют право на существование. При чтении экологических работ нужно иметь в виду, что многие исследователи не выделяют эти подходы и только из контекста можно понять (далеко не всегда), о чем идет речь. А.А. Ковальчук в каждом сообществе инфузорий выделял вид-доминант, имевший наиболее высокую численность, и вид-лидер с наибольшим энергобюджетом. При совпадении вида-доминанта с видом-лидером сообщество получало название “одноименного” (45). Необходимо также остановиться на связи термина “доминант” с терминами “эдификатор” и “детерминант”, понимаемыми тоже по-разному, иногда определения этих терминов весьма расплывчаты, например,
А.А. Шенников называет эдификаторами доминанты, которые обуславливают особенности данного ценоза, а Б. И. Миркин и Г. С. Розенберг - виды, контролирующие режим отношений в сообществе (70). Эдификаторами называют и центральные члены консорций (7, 109), иногда между эдификаторами и детерминантами не делают различия (17, 109). Но наиболее часто термин “эдификатор” употребляется для обозначения организмов, в наибольшей степени изменяющих окружающую среду (17, 44, 67, 92, 115). В. Б. Миркин (65) называет детерминантами виды, несущие максимум информации об условиях среды, но чаще этим термином обозначают виды, наиболее сильно влияющие на другие виды. Предложено оценивать детерминирующее действие вида по тем изменениям, которые наступают в сообществе после удаления из него данного вида (137).Подобный разнобой в употреблении терминов создает большие трудности для понимания экологической литературы, особенно в тех случаях, когда, например, гидробиолог или зоолог знакомится с фитоценологическими работами. Желательно унифицировать употребление этих терминов, хотя из истории экологии известно, что такие попытки имеют мало шансов на успех в силу приверженности исследователей к привычной им терминологии. Можно предложить следующую трактовку разбираемых понятий, основанную на наиболее часто используемых в экологической литературе их значениях. Доминант (в узком смысле) - вид, имеющий наибольшую величину показателя количественной представленности в изучаемом сообществе. Этим показателем может быть в зависимости от цели исследования численность, биомасса, продукция, проективное покрытие, встречаемость... или какая-то их комбинация. Эдификатор - вид с наиболее сильно выраженной средообразующей способностью, т.е. производящей наибольшие изменения в среде. Эдификаторами могут быть, например, центральные организмы консорций, в водоеме - друзы моллюсков дрейссен, которые значительно изменяют условия на биотопе, гнилостные и десульфирующие бактерии, благодаря жизнедеятельности которых в воде накапливается сероводород, метанообразующие бактерии и т. п. Детерминант - вид, оказывающий наибольшее влияние на другие виды сообщества. Сюда относятся и “ключевые виды” биоценозов, чье влияние значительно превосходит их долю в численности или биомассе всех организмов ценоза. Часто ключевые виды - хищники, занимающие высокое место в пищевой пирамиде и потребляющие организмы, которые без сдерживающего влияния хищника могли бы монополизировать пространство или пищевые ресурсы и вытеснить менее конкурентноспособные виды (Несис К. Н.) (81). Разумеется, один и тот же вид может является одновременно доминантном, детерминантом и эдификатором, но так бывает не всегда. Когда исследователи не различают значения этих терминов, они говорят о доминировании в широком смысле слова. За термином “индикатор” сохраняется значение - вид, характеризующий условия среды.
Между экологами ведутся споры, выделять ли доминанты для всего сообщества, включающего растительные и животные организмы, или же выделять их отдельно для автотрофов и гетеротрофов или даже внутри более мелких подразделений. Например, полагая, что доминирование - присвоение пространства потенциальной экологической ниши некоторого подчиненного вида другим видом, что в явной форме проявляется внутри одного трофического уровня, делается вывод, что продуцент не может доминировать над консументом или декомпозитором, поскольку непосредственные источники получения ими энергии не перекрываются (149). Здесь доминирование связывается с конкурентными взаимоотношениями, но конкуренция может быть не только за пищу но и за другие условия обитания, например, за место. Доминирование в предложенном нами понимании можно оценивать как для всего сообщества, так и для любой его части. Эдификаторов и детерминантов можно выделять лишь для той группы организмов, внутри которой проявляется соответствующий эффект.
До сих пор в качестве элементов системы, о доминировании которых судят, брали вид (популяцию, ценопопуляцию), но доминирование может быть и внутривидовое. Тогда в качестве элемента системы выступают различные половые, возрастные, физиологические группировки внутри вида. Например, известно, что в водоеме весной после вскрытия льда среди хирономид доминируют личинки IY возраста, а после вылета имаго и откладки яиц начинают доминировать личинки I возраста. В популяциях рыб также наблюдается доминирование отдельных возрастных групп (83). Некоторые возрастные стадии животных могут выступать в качестве детерминантов, подавляющих развитие младших возрастов. Такой эффект особенно хорошо изучен на примере развития лягушки (116), личинки III личиночного возраста майского жука подавляют развитие личинок I и II возрастов (26).
Иногда доминанты выделить трудно, нередко в растительных сообществах фитомасса примерно поровну распределяется между 5 - 10 основными строителями сообщества. В этом случае можно говорить о коллективном доминировании и выделять группы доминирования. К одной группе обычно относят растения одной жизненной формы и сходной экологии (69, 70). В качестве доминантов могут быть выступать таксоны и надвидового ранга (114,115); например, в сухих травянистых степях часто доминируют саранчевые (112). В качестве единиц группового (коллективного) доминирования могут выступать любые биоценотические единицы: синузии (82), эргоцены, гильдии и т. п. В.П. Воробьев (22) говорит о доминировании в бентосе Азовского моря одного биоценоза над другим, причем во всех биоценозах наблюдается чередование интенсивности развития представителей эпифауны и инфауны, например, в биоценозе митилястера I и II места занимают организмы эпифауны, II
I и IY - инфауны, Y и YI - снова эпифауны, затем инфауны и т. д.Терминология, применяемая для описания различных степеней доминирования, чрезвычайно запутана, особенно фитоценологами, о чем Н.В. Тимофеев-Ресовский (99. стр. 43) сказал так: “Я не фитоценолог, но мне приходится иметь дело с фитоценологическим материалом. Это каторжная работа. Чтобы понять короткую статью, нужно сначала разобраться в словоупотреблениях. Представитель одной школы употребляет слово в одном значении, другой ученый, употребляя это же слово, описывает, оказывается, совершенно другие процессы. Практически почти во всех биохорологических дисциплинах терминология совершенно произвольна, и это делает огромный накопленный материал почти несравнимым. Количество, так сказать, не переходит в качество. Материала много, а с ним ничего не сделаешь, невозможно даже перевести все это на какой-то один язык”.
Степень доминирования вида в фитоценозе часто оценивается по величине проективного покрытия - сумме проекций наземных частей растений этого вида в процентах от площади пробы. Иногда используется удельное проективное покрытие - процент покрытия данного вида от общего покрытия всего сообщества. Для количественной оценки используются шкалы проективного покрытия, % :
В.С. Ипатов и др. (41) считают, что удельное проективное покрытие точнее выражает степень участия видов в сложении растительного покрова, т.е. несет большую фитоценотическую нагрузку. Они использовали следующую шкалу: господствующие виды - удельное покрытие более 66% , согосподствующие - 33-36, наполнители 5-33, редкие виды - менее 5%. Когда величина проективного покрытия близка к границам классов, допускается промежуточная оценка: вид господствует-согосподствует - 49-83, согосподствует - наполнитель - 19-48, наполнитель - редкий – 2.5-18%. Для воспроизводимости оценок проективного покрытия считается достаточной доверительная вероятность 80%. Одна из новейших и наиболее обоснованных - шкала Е.Л. Любарского (59), названная им “КТЩ-5”, т.е. 5-балльная квадратично-трансформированная шкала, которая “обеспечивает математически закономерное нарастание линейной неравномерности с учетом двумерной сущности покрытия, проецируемого на одномерную шкалу”: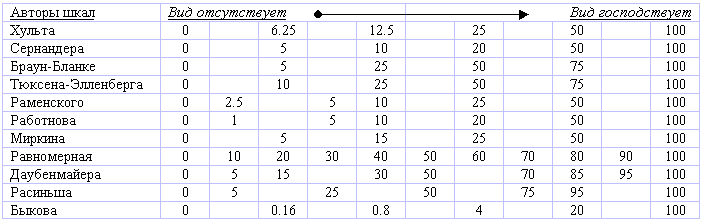
|
Балл |
Границы классов покрытия, % |
Ценобионтное значение |
|
0 |
0 |
Отсутствие |
|
1 |
0-4 |
Наличие |
|
2 |
4-16 |
Соучастие |
|
3 |
16-36 |
Согосподство |
|
4 |
36-64 |
Господство |
|
5 |
64-100 |
Абсолютное господство |
При невозможности получить надежные количественные показатели используют словесную оценку обилия с использованием различных шкал: Тенсли, Браун-Бланке, Хенсона, Друде (17).
Характер доминирования деревьев может оцениватся по шкале Крафта (17): 1 балл - исключительно господствующие, выделяются над пологом древесины, 2 - господствующие, главная часть древесного полога, 3- согосподствующие, входят в общий полог, но затенены, 4 - угнетенные, достигают полога древесных крон, 5 - вполне угнетенные, почти лишенные ветвей, отмирающие.
При характеристике роли вида по обилию используются также шкала Крогеруса (58): доминанты - составляют более 5% общего количества особей, инфлюенты - от 2 до 5%, рецеденты - менее 2% . В гидробиологии широко применяются четыре градации доминирования (47, 72): доминанты. субдоминанты, второстепенные и случайные виды.
При учете организмов методом выборочного обследования их численность (при прочих равных условиях) пропорциональна количеству собранных проб, т. е., обследованной площади, поэтому для выделения классов доминирования по численности может использоваться шкала Любарского, а для биомассы можно предложить аналогичную кубически трансформированную шкалу, поскольку биомасса равна численности, умноженной на средний вес особи.
Шкала доминирования по численности и биомассе
|
Балл |
Границы классов по численности |
Границы классов по биомассе |
Название степени доминирования |
|
1 |
0 < N £ 4 |
0 < B £ 1 |
Малозначимый вид |
|
2 |
4 < N £ 16 |
1 < B £ 6 |
Второстепенный |
|
3 |
16 < N £ 36 |
6 < B £ 22 |
Cубдоминант |
|
4 |
36 < N £ 64 |
22 < B £ 50 |
Доминант |
|
5 |
64 < N £ 100 |
50 < B £ 100 |
Абсолютный доминант |
Здесь
N - доля вида в общей численности, % ; B - в общей биомассы, %.Принципы построения различных шкал доминирования и описание некоторых более современных и сложных шкал можно найти в работах ((30, 31, 86).
В разных пробах из одной выборки могут доминировать различные виды, большую роль при этом играет случайность, поэтому для установления закономерностей доминирования нужно рассматривать весь материал выборки, характеризующий определенный участок местности. При этом можно заметить, что популяция одного вида, доминирующая в определенном биотопе, в других биотопах может играть второстепенную роль. Одни виды встречаются в качестве доминантов часто, другие редко или вообще не встречаются. Характер доминирования, его устойчивость во времени и пространстве у разных видов неодинаковы, поэтому недостаточно просто установить факт доминирования. Возникает необходимость в разработке типологии
доминантов. По способности быть доминантом организмы относятся к определенному ценотипу. Ценотип - это группы видовых популяций (популяционный ценотип) или видов (видовой ценотип), которые характеризуются сходной ролью (или потенциальной способностью играть определенную роль) и ценотической позицией в пределах биоценоза или региона ((17, 70). Особенно давно типология ценотипов разрабатывается в фитоценологии, при этом используется ограниченный набор свойств, не всегда наиболее существенных. Поэтому Х.Х. Трасс (101,102) предложил набор признаков, наиболее существенных для выделения ценотипов:1. Площадь, покрываемая особью доминирующего вида. Отсюда доминанты делятся на нано- , микро-, мезо-, макро- и мегадоминанты.
2. Величина создаваемого доминантами участка (заросли, группировки). Отсюда деление на нано-, микро-, мезо-, макро-, и мегагрегидные доминанты. Для животных это величина стада, стаи...
3. Различная сила влияния на другие компоненты ценоза и на биотоп, т. е. способность организмов быть эдификаторами и детерминантами. Эдификаторы делятся на гипер-, суб- и гипедификаторы.
4. Доминантный состав фитоценоза в целом. Ценозы делятся на монодоминантные, когда доминируют один вид, полидоминантные - несколько видов имеют сходные показатели доминирования и в этом случае называются содоминантами. Среди доминантов можно различать такие, которые всегда образуют чистые заросли - солодоминанты. Виды, образующие агрегации с двумя - тремя содоминантами называются кондоминантами, а виды, встречающееся совместно с многочисленными слабодоминирующими видами - миксодоминантами.
5. Встречаемость вида в роли доминанта. Облигатные доминанты - виды, которые встречаются всегда или преимущественно как доминанты. Часть ареала вида, в котором он является доминантом, называют “ареалом доминирования”. Факультативные доминанты встречаются как в качестве доминантов, так и в качестве сопутствующих видов. Адвентивные доминанты (преваленты) - виды, доминирующие случайно, при особенно благоприятных для них условиях среды. Отсюда, например, вытекает метод выделения культур микроорганизмов на специфических питательных средах.
6. Сезонность доминантов. Консерводоминанты доминируют весь вегетационный период, темпоральдоминанты - в определенное время года.
Наиболее известны следующие классификации ценотипов (16, 17, 70, 101):
Браун-Бланке и Павийяр выделяли популяции строителей (эдификаторов), консерваторов (поддерживающих), консолидаторов (укрепляющих), нейтральных видов, деструкторов (разрушающих). И.К. Пачосский различал превалиды, компоненты и ингредиенты. Г.И. Поплавская и В.Н. Сукачев выделяли эдификаторы (популяции, способные быть строителями основных ярусов фитоценоза), соэдификаторы и ассектаторы (мало влияющие на среду фитоценоза). По роли в динамике сообщества эдификаторы подразделялись на аутохтонные и дегрессивные, по характеру взаимодействий ассектаторы и соэдификаторы могут быть эдификаторофилы и эдификаторофобы. С.Я. Соколов внутри каждой градации предыдущих авторов по тенденции поведения в сукцессионном процессе выделил прогрессивные, регрессивные и устойчивые популяции, а также автохтонные и антропогенные виды. Е.М. Лавренко различал эдификаторы, превалиды, субпревалиды и ассектаторы. Г.И. Дохман за основу классификации взял два показателя - обилие и проективное покрытие. Он выделял доминанты 1 порядка - высокое обилие и высокое покрытие, доминанты II порядка - высокое обилие при среднем покрытии, доминанты III порядка- высокое обилие при низком покрытии. А.Б. Уильямс различал первичные доминанты, вторичные доминанты, случайные доминанты и субдоминанты.
Клементс выделял: I. Климаксовые доминанты, включающие пердоминанты, эудоминанты, кодоминанты и субдоминанты. II. Сериальные доминанты. III. Доминанты микрогруппировок, включающие доминулы, субдоминулы и эвдоминулы. IY. Второстепенные виды. Инфлюенты (влияющие виды) по степени влияния он подразделил на перфлюенты, эуфлюенты, субфлюенты и вефлюенты.
Развернутую подробную классификацию видовых и популяционных фитоценотипов предложил также Ю.Р. Шеляг-Сосонко. Б.А. Быков (17) разработал общую классификацию для растений, животных и микроорганизмов. Доминанты, субдоминанты и кондоминанты он делит на патулекторы, дензекторы, коннекторы, терректоры, предоминанты, гифекторы, микробекторы, сестонекторы. Выделив эзодоминанты, он делит их на консорбенты и доминуленты. Ингредиенты же подразделяются на ассектаторы и адвентикаторы. Видно, что одни и те же термины, входя в разные классификации, получают неодинаковые значения.
Наиболее просто получаемый показатель значимости вида в сообществе - его встречаемость. При этом различают абсолютную встречаемость - отношение количества проб, в которых обнаружен данный вид, к общему количеству проб, и относительную встречаемость - отношение абсолютной встречаемости к сумме встречаемостей всех видов биоценоза. Б.А. Вайнштейн (19) предлагал заменить термин встречаемость термином разброс, но это не прижилось. поскольку разброс используется в другом значении для характеристики размещения организмов в пространстве. Но предложение этого исследователя обозначить встречаемость во времени термином “постоянство” заслуживает внимания. Количественное выражение встречаемости зависит от величины пробы и характера статистического распределения особей по биотопу. На встречаемость оказывают большое влияние миграции организмов, в связи с чем К. Фридерикс ввел ряд терминов (112). Животных которые в течении всей жизни обитают в одном биоценозе, он назвал гомоценными, а которые с известной правильностью меняют биоценозы - гетероценными. В последнем значении сейчас чаще употребляют термин “гетеротопные”, который не нужно путать с термином “эвритопные”, т.е. обитающие в разных биоценозах (на разных биотопах).
Организмы, встречающиеся только на данном биотопе или находящиеся здесь всегда в большем числе, чем другие, он называл руководящими формами (это хорошие индикаторы биотопа). Растения, встречающиеся на каком-то участке чаще, чем на других, ботаники преферентами, а виды, которые встречаются не каждый год - случайными. Виды, которые хотя и встречаются постоянно на данном биотопе, но в меньшем количестве, чем на других биотопах, он назвал отступающими. Виды приходящие в биотоп изредка, например, за пищей, водой... называются посетителями. Наконец, заблудившиеся виды - хотя и держатся в данном биотопе некоторое время, но не чувствуют себя здесь как дома.
В гидробиологии наиболее часто употребляется следующая шкала встречаемости (22, 39, 40): константные виды - встречаемость более 50%, второстепенные - 25-50%, случайные - менее 25%. Ботаники константными называют виды с встречаемостью 80-100%, а виды встречаемостью более 50% - постоянными (17). При изучении фитопланктона применялась другая шкала (46): наиболее часто встречающиеся виды - более 50%, часто встречающиеся - 20-50%, нечасто встречающиеся - 1-20%, редко встречающиеся - менее 1%. Поскольку встречаемость, как и покрытие, связана с площадью, то для нее может быть пригодна шкала Любарского.
При вычислении различных экологических показателей, в которые входит величина встречаемости или число видов, необходимо помнить, что эти параметры зависят от величины обследованной площади (количества проб, объема собранного материала). При увеличении площади сбора число обнаруженных видов растет, затем рост замедляется по мере приближения к некоторой величине, равной полному числу видов на обследуемом участке. Впервые подобная закономерность была отмечена французским исследователем Жаккаром в 1902 г., графическое изображение ее показано на рисунке 1.
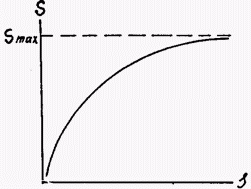
Рис. 1. Теоретическая зависимость числа обнаруженных видов (S) от величины обследованной площади
(s). Smax – полное число видов на участке.Для математического описания связи виды - площадь чаще всего привлекается степная функция
S
= a · nb ,где
S - число обнаруженных видов, n - количество проб, a и b - константы. (123). В качестве примера мы взяли число видов хирономид, найденных нами в бентосе оз. Плещеево (Ярославская обл.) осенью 1979 г. Пробы отбирались дночерпателем с площадью захвата 1/40 м2. Следует отметить, что личинки хирономид, как и многие другие группы гидробионтов, далеко не всегда могут быть определены до вида, иногда их идентифицируют только до группы видов или до личной формы. Это cказывается только на константах в уравнении. Поэтому, когда гидробиологи употребляют термин “вид”, часто он обозначает низший таксон, до которого можно определить найденный организм по существующим определителям. Для хирономид оз. Плещеево аппроксимирующая функция имела вид S = 4.33 n0,39, коэффициент детерминации r2 = 0.88. Некоторые исследователи интерпретируют параметр b как меру пространственного однообразия населения, хотя это не всегда так (86). Все более точно наш эмпирический материал соответствовал формуле Глисона (132)S
= a + b log n,при этом
a = 3.92, b = 3.48, r2 = 0.94. Подобная зависимость соответствует гипотезе Вильямса (164) о том, что увеличение числа видов является следствием увеличения количества разных типов местообитаний при возрастании обследуемой площади. Сходными уравнениями описывается также увеличение частоты встречаемости при увеличении площади проб и связь количества видов с числом собранных особей. Но обе вышеприведенные формулы непригодны для отыскания полного количества видов, поскольку они не асимптотические, т.е. при увеличении площади сбора количество видов стремится к бесконечности, что не соответствует действительности. Для решения этой задачи мы предлагаем воспользоваться уравнением экспоненциального роста, нашедшим широкое применение при описании роста животных (в литературе оно называется также формулой Броди или уравнением Берталанфи)S
= A - B e-cn,где
B и C - константы, A - асимптота, показывающая в данном случае сколько видов можно обнаружить на изучаемом участке в данный момент времени при использовании данной методики сбора. В нашем случае оно имеет вид S= 20 – 13.2 e – 0.28n (рис. 2). Методика нахождения параметров уравнения изложена в работах (63, стр. 84-85; 64, стр. 27).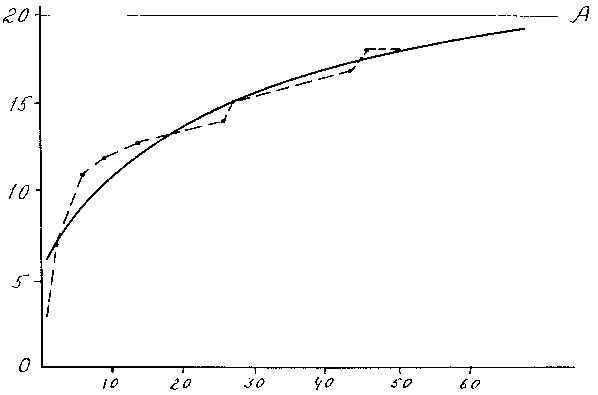
Рис. 2. Зависимость числа видов хирономид от количества проб
Сплошная линия – теоретическая кривая, штриховая линия – эмпирические данные.
Рассматривая зависимость количества видов от числа особей, А.П. Левич нашел, что у планктонных организмов в любой выборке существуют виды единичной численности, т.е. рассматривая эту функцию, можно прийти к выводу, что она строго возрастает. Это свойство он назвал “правилом экологической неаддитивности”. Благодаря ему не существует естественного сообщества, являющегося просто суммой отдельных частей. Вместе с ростом численности в сумме должны появится новые виды. В монографии (56) приведены формулы зависимости количества видов от числа особей для разных типов распределения.
При рассмотрении вопроса о зависимости числа видов от их встречаемости обычно используют равномерную шкалу встречаемости, разделенную на 5 классов с интервалом 20%. Тогда форма этой связи графически изображается кривыми трех типов (рис. 3).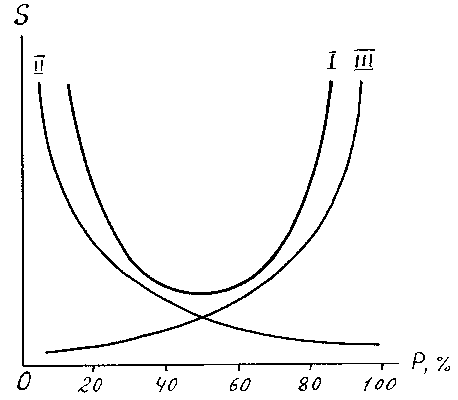
Рис. 3. Теоретическая связь числа видов (
S) с их встречаемостью (P).В практике экологических исследований часто встречаются кривая I типа, т. е.
U -образная, называемая иногда кривой Раункиера (70). Обычно она наблюдается при некоторой определенной величине пробы в пределах относительно однообразного по распределению видов (гомотонного) сообщества (6, 18, 19). В этом случае большинство видов обладают либо незначительной, либо высокой встречаемостью, а видов с встречаемостью 40 - 60% очень мало. Считается, что это говорит о удачном подборе размера пробы (учетной площадки). Если же большинство видов встречается во всех пробах (кривая III типа), то размер пробы слишком велик, если встречаемость большинства видов низка (кривая II типа), то размер пробы мал - ситуация, часто встречающаяся в гидробиологии. Изучая видовой состав зоопланктона в зарослях высшей водной растительности, Л.Н. Зимбалевская (36) обнаружила, что в ассоциациях разных видов макрофитов наблюдаются все типы этой зависимости. Распределение хирономид в оз. Плещеево имеет форму кривой II типа (рис. 4), что свидетельствует как о гетерогенности этого водоема, так и о недостаточном размере пробы для достаточно полного учета фауны хирономид.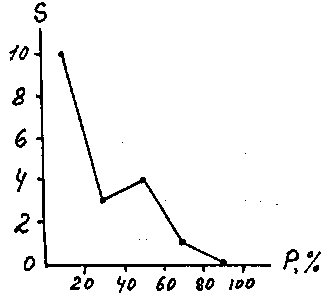
Рис. 4. Связь числа видов хирономид с их встречаемостью в оз. Плещеево
II. КОЛИЧЕСТВЕННАЯ ОЦЕНКА СТЕПЕНИ ДОМИНИРОВАНИЯ.
Когда говорят о количественных оценках доминирования, то обычно понимают этот термин в узком смысле, т.е. как количественную представленность в сообществе. Оценка степени эдификаторного или детерминирующего воздействий производится редко и чаще всего в эксперименте. Для нее нужны специфические методы. Выше уже упоминалось предложение (137) учитывать воздействие одних видов на другие по величине изменения продуктивности сообщества в результате удаления испытуемого вида. Для этой же цели используется метод парциальных сопряженностей: рассчитывают сопряженность между видами А и Б в присутствии и в отсутствии вида С. Если парциальные сопряженности достоверно не различаются, можно считать, что эти виды индифферентны к виду С. Таким путем было показано, например, слабое влияние луговых доминантов на состав второстепенных видов. В экспериментах изучают поведение видов в чистом посеве и в смеси, находят эмпирические коэффициенты конкуренции, по величине которых ранжируют виды и судят о степени их влияния на другие виды (69).
Оригинальный метод выявления видов, оказывающих наибольшее влияние на другие, предложил В.Д. Федоров (106). Он считает, что, если виды, составляющие сообщество, развиваются независимо друг от друга, т.е. отсутствует конкуренция за источники питания и пространство, то в этом случае каждый из них имеет равные возможности для развития и, следовательно, равную вероятность в данный момент времени иметь ту численность, которая определяется его кривой роста. Отсюда следует, что в отобранной пробе любой вид имеет одинаковую вероятность быть наиболее редким или же наиболее обычным. Следовательно, распределение численности по видам должно подчинятся нормальному закону. Им предложен метод построения графика, на котором теоретические значения численности лежат на прямой линии. Если точка, характеризующая численность определенного вида, достоверно отклоняется вправо от этой прямой, значит этот вид угнетает те виды, которые лежат левее прямой. Следует отметить, что допущения, лежащие в основе этого метода, нельзя считать вполне обоснованными. Кроме того, этот метод достаточно сложен для расчетов, в самой же работе он изложен столь конспективно, что не дает возможности им пользоваться. Поэтому на практике он применяется очень редко (29).
Для количественной характеристики какого-либо явления или процесса (в данном случае доминирования) надо построить понятие особого рода - показатель. Он должен давать количественную характеристику явления в единстве с его качественной определенностью. Показатель состоит из модели количественной стороны явления, устанавливающей, что, где, когда и каким образом подлежит измерению (качественный реквизит), и числового выражения величины явления в конкретных условиях места и времени (количественный реквизит) (120). Построение показателя является развитием экологического понятия с точки зрения его количественной оценки. Как правило, каждому экологическому понятию должен соответствовать определенный показатель, ибо вводить в науку понятия, которые нельзя измерить, мало целесообразно. Известно, что всякое явление или процесс могут иметь очень много различных сторон или аспектов. Лишь сравнительно простое явление может характеризоваться одним простым показателем. Сложные явления приходится характеризовать целой системой показателей, каждый из которых измеряет какой-то один аспект явления. Удобнее в таком случае использовать сложные, комплексные, интегральные показатели, объединяющие ряд простых. При этом отдельным сторонам явления может придаваться разное значение, поэтому характеризующие их показатели входят в интегральный показатель с разными коэффициентами (весами), отражающими роль данной стороны явления в общей его оценке. Например, Браун-Бланке (цит. по 58) при изучении структуры растительности ввел “коэффициент обилия - доминантности”. При этом он полагал, что покрытие играет более существенную роль, чем обилие, в тех случаях, когда степень его высока, в то же время как обилие становится определяющим в противоположном случае. Иногда для одного и того же понятия различными исследователями строятся разные показатели, носящие одинаковое название, но характеризующие явление с разных сторон. В этом случае бессмысленно выяснять, какой показатель “правильнее”.
При изучении доминирования нужно различать два аспекта. Во-первых, нужно установить, какой вид доминирует в сообществе и оценить степень его доминирования. Отражающий эту сторону доминирования показатель мы назовем индексом индивидуального доминирования. Во-вторых, нужно отразить характер доминирования в сообществе в целом, чтобы иметь возможность сравнивать по особенностям доминирования разные сообщества. Для этого нужно оценить упорядоченность видов по индексам индивидуального доминирования, соответствующие показатели мы назовем индексами структуры доминирования. Индексы могут быть чисто эмпирические, или же основываться на каких-то теоретических соображениях. При разработке индексов желательно одновременно предлагать способ оценки их точности, т.е. формулу стандартной ошибки индексов. Это даст возможность с помощью статистических методик оценивать достоверность разницы между индексами. Необходимо также выяснить, имеет ли показатель систематическую ошибку, т.е. постоянное завышение или занижение измеряемой величины. Нужно знать, как реагирует показатель на изменение величины учетной площадки, объема выборки, общего объема собранного материала, зависит ли он от типа статистического распределения изучаемых организмов. Необходимо выяснить чувствительность или устойчивость показателя, т.е. как он реагирует на ошибки в исходных данных и остаются ли колебания его величины при наличии таких ошибок в достаточно ограниченных и практически допустимых пределах. К сожалению, для существующих индексов доминирования такая работа практически не проводилась.
В зависимости от того. используется для вычисления индексов один параметр (численность, биомасса, встречаемость...) или несколько, все индексы можно разделить на одно-, двух- и многопараметрические.
А. Индексы индивидуального доминирования.
Наиболее употребительным индексом такого рода является доля (процент), которую составляет показатель обилия вида (численность, биомасса, продукция, покрытие...) от общего обилия всех видов в выборке из сообщества. Например, в большинстве гидробиологических работ, когда говорят о доминирующем виде без указания способа определения доминирования, имеется в виду просто наиболее обильный вид. Часто к доминирующим видам относятся виды, занимающие по обилию I, II, III места. Э3кологические ниши видов могут быть настолько сближены, что преобладание того или иного вида зависит часто от случайных моментов. Поэтому придавать большое значение незначительным различиям в степени доминирования не следует (66). При учете численности хирономид оз. Плещеево, например, (рис. 5) стандартные ошибки учета оказались от 64% у первого вида, до 19.1% у вида № 5. В гидробиологических полевых исследованиях стандартные ошибки равняются обычно несколькими десятками процентов, превышая иногда 100%. В фитоценологии при сравнении индексов доминирования иногда используют не величину их стандартной ошибки, а, исходя из задачи исследования, заранее устанавливают величину минимально необходимого превышения, которая дает основание для оценки определенной группы растительности как преобладающей (68).
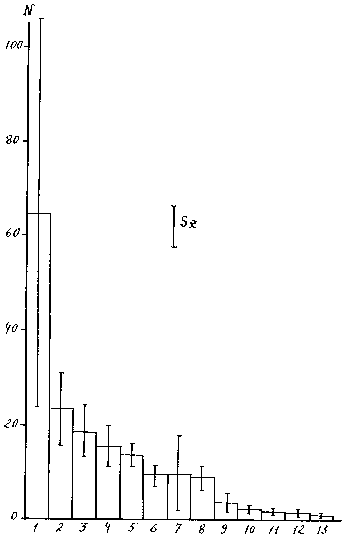
Рис. 5. Распределение отдельных видов хирономид оз. Плещеево по численности.
По оси ординат – численность, экз./пробу; по оси абсцисс – порядковый номер вида,
В фитоценологии для характеристики доминирования применяют показатель называемый “частота доминирования”,
DF, введенный Де Фризом (12, 61, 129). Он равен количеству проб, в которых данный вид занимал I -III места (иногда только 1 место), выраженному в процентах от общего числа проб. О.М. Кожова (46) по величине этого индекса делила виды фитопланктона Братского водохранилища на 3 категории:Оригинальный метод оценки доминирования предложил Сандерс (158). В каждой пробе выделяются первые десять по численности (или иным показателем обилия) видов. Первому присваивается вес - 10, второму 9, десятому - 1, затем для каждого вида находится величина
d
= S wini,где
wi -вес i - го места, ni - количество проб, в которых вид занял i - место. Чем больше величина индекса, тем сильнее доминирование данного вида (формула и обозначения наши). Но поскольку величина индекса зависит от общего количества проб, то для возможности сравнения разных выборок мы предлагаем найденную сумму делить на количество проб, d’ = S wini / n. Тогда величина индекса будет меняться от 0 - отсутствие вида, до 10 - доминирование во всех пробах. Величина этого индекса говорит о “силе доминирования”, но из этих же данных можно вывести показатель. который можно назвать “устойчивость доминирования”, показывающий, насколько меняется величина доминирования в разных пробах. Это покажет коэффициент вариации величины wini относительно ее среднего значения, равного 0.1S Wini. Применение этого метода к материалам по хирономидам оз. Плещеево показало, что индекс доминирования Сандерса сильно коррелирует с показателем встречаемости, чаще встречающиеся виды обнаруживают и более сильное доминирование. Но чаще встречающие виды - не обязательно самые обильные, например, самый обильный вид Paratanytarsus sp. показал невысокую встречаемость (18%) и слабое доминирование (d¢ = 1.4). Наименее устойчивое доминирование, т. е. наиболее высокие коэффициенты вариации, показали виды с высокими и низкими индексами доминирования. Наибольшая устойчивость доминирования отмечена у видов со средней силой доминирования. Разновидностью этого метода является оценка доминирования по так называемому “биологическому индексу” (147).Если в распоряжении исследователя имеется несколько выборок, то виды внутри каждой выборки можно ранжировать по величине индекса доминирования, затем проверить сопряженность рангов с помощью, например,
W - статистики Кендала. Доверенность сопряженности говорит об устойчивом характере доминирования (29, 130). Для графического изображения доминирования чаще всего примеряют гистограммы, подобные изображенной на рис. 5, но есть и другие приемы. Романишин (цит. по 138) предложил изображать доминирование прямоугольником, площадь которого пропорциональна величине индекса (рис. 6). Римскими цифрами обозначаются выборки, характеризующие биотоп или сезон года, арабскими - виды.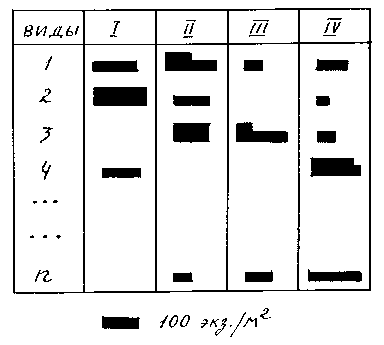
Рис. 6. Изображение доминирования по методу Романишина.
Похожий график применил В.Д. Федоров для изображения доминирования отдельных видов в фитопланктоне Белого моря (106). Он выделил три степени доминирования (не указывая, каким образом), каждая степень обозначалась определенной штриховкой (рис. 7).
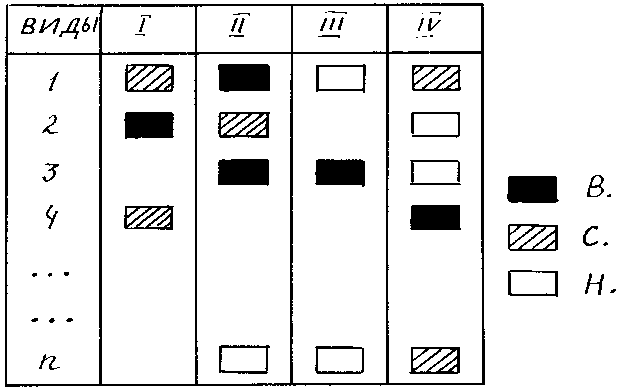
Рис. 7. Изображение доминирования по методу Федорова
В – высокий, С – средний, Н – низкий уровни доминирования
Более сложно характеризовать доминирование сразу по нескольким параметрам, например, биомассе (B), численности (N), встречаемости (P) и т. д., т.е. D = f(B, N, P,...). В.И. Василевич (20) считает нецелесообразным заменять этот набор величин каким-то одним комплексным показателем. Поскольку с точки зрения математики этот набор можно рассматривать как вектор (упорядоченная система чисел), то он предлагает роль вида в сообществе характеризовать вектором, компонентами которого служат характеристики обилия, распределения и жизненности вида. Например, вектор А (20, 35, 17, 3) означает: покрытие - 20, вес на единицу площади - 35, среднеквадратическое отклонение покрытия - 17, жизненность - 3 балла. Для обработки таких описаний может быть применен аппарат высшей алгебры. На практике подобный метод не нашел широкого применения - исследователи предпочитают строить комбинированные показатели доминирования.
В гидробиологии такие показатели чаще всего представляют комбинацию 2-3 параметров: B, N, P. Особенно широкое применение нашли различные модификации так называемого “индекса плотности” Бродской - Зенкеевича, который стал рассматриваться как индекс доминирования. Первоначально он был предложен в форме 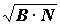 , причем эти величины брались в абсолютных значениях (34). По величине индексов все виды делились на руководящие, характерные I порядка, характерные II порядка, второстепенные I порядка и второстепенные II порядка. Были построены графики значений этих индексов для разных видов, расположенных в порядке убывания величины индексов, отмечалось, что графики имеют ступенчатый характер и не пересекаются. Такую форму индекса применяли многие исследователи (27, 28, 39, 47), иногда он рассчитывался без извлечения корня (43). В дальнейшем В.А. Бродская и Л.А. Зенкевич индекс использовали в форме
, причем эти величины брались в абсолютных значениях (34). По величине индексов все виды делились на руководящие, характерные I порядка, характерные II порядка, второстепенные I порядка и второстепенные II порядка. Были построены графики значений этих индексов для разных видов, расположенных в порядке убывания величины индексов, отмечалось, что графики имеют ступенчатый характер и не пересекаются. Такую форму индекса применяли многие исследователи (27, 28, 39, 47), иногда он рассчитывался без извлечения корня (43). В дальнейшем В.А. Бродская и Л.А. Зенкевич индекс использовали в форме 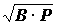 , ставшей наиболее распространенной (13), причем биомасса бралась в абсолютных единицах, а встречаемость в процентах. Эта работа и сейчас находится в сфере активного цитирования. Но практически этот индекс был впервые опубликован в работе В.Г. Богорова (11) еще в 1934 г., он использовал его для оценки степени доминирования различных компонентов в пище сельди, причем ссылался на работу В.А. Бродской Л.А. Зенкевича, которую видел, очевидно в рукописи, поскольку вышла она значительно позднее. Он рекомендовал брать величину биомассы не в абсолютных единицах, а также в процентах. Индекс доминирования в форме использовали многие гидробиологи (22, 47, 71-74, 88, 100), А А.Г. Наумов (76) использовал сразу обе формулы - и . Специально рассматривался вопрос (27, 28), какой из этих формул отдать предпочтение. Н.Е. Денисов считал, что биомасса может служить мерой преуспевания популяции в олиготрофных глубинах океана, где в основе отношений между организмами лежит конкуренция за пищу. Для мелководной же зоны характерна обостренная конкуренция за жизненное пространство, поэтому показатель состояния популяции в этом случае должен зависеть как от биомассы, так и от численности, так как при одинаковой биомассе большая площадь биотопа контролируется популяцией, имеющей большую численность, поэтому он рекомендовал формулу . Но B = Nb, где b - средний вес организма. Значит рекомендованную Н.Е. Денисовым формулу можно записать в виде
, ставшей наиболее распространенной (13), причем биомасса бралась в абсолютных единицах, а встречаемость в процентах. Эта работа и сейчас находится в сфере активного цитирования. Но практически этот индекс был впервые опубликован в работе В.Г. Богорова (11) еще в 1934 г., он использовал его для оценки степени доминирования различных компонентов в пище сельди, причем ссылался на работу В.А. Бродской Л.А. Зенкевича, которую видел, очевидно в рукописи, поскольку вышла она значительно позднее. Он рекомендовал брать величину биомассы не в абсолютных единицах, а также в процентах. Индекс доминирования в форме использовали многие гидробиологи (22, 47, 71-74, 88, 100), А А.Г. Наумов (76) использовал сразу обе формулы - и . Специально рассматривался вопрос (27, 28), какой из этих формул отдать предпочтение. Н.Е. Денисов считал, что биомасса может служить мерой преуспевания популяции в олиготрофных глубинах океана, где в основе отношений между организмами лежит конкуренция за пищу. Для мелководной же зоны характерна обостренная конкуренция за жизненное пространство, поэтому показатель состояния популяции в этом случае должен зависеть как от биомассы, так и от численности, так как при одинаковой биомассе большая площадь биотопа контролируется популяцией, имеющей большую численность, поэтому он рекомендовал формулу . Но B = Nb, где b - средний вес организма. Значит рекомендованную Н.Е. Денисовым формулу можно записать в виде 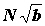 , т.е. в ней численности придается значительно большее значение, чем среднему весу, что далеко не всегда оправдано.
, т.е. в ней численности придается значительно большее значение, чем среднему весу, что далеко не всегда оправдано.
Придавая различную значимость биомассе и численности и беря их в относительных единицах, была предложена формула (51), которая при определенных допущениях может оценивать роль вида в потоке энергии:
Pi
= Bi0.75 Ni0.25 / S (Bi0.75 Ni0.25).В исследованиях морских экосистем индекс
стал особенно широко использоваться после классической работы В.П. Воробьева (22), в которой он по величине этого индекса делил виды на руководящие, характерные I и II порядка, второстепенные и случайные, четких объективных критериев отнесения видов к этим группам он не дал. Руководящие и характерные виды (а в другом случае только руководящие) он относил к ядру биоценоза. Биомасса бентоса колеблется в очень широких пределах - от нескольких миллиграмм до сотен грамм в пробе, тогда как встречаемость изменяется от 0 до 100%, это может привести к завышению значения биомассы. Поэтому были рекомендованы формулы 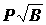 или
или 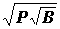 , что уменьшает значение биомассы (73, 74). По этим индексам виды делились на доминантные (руководящие), субдоминантные и второстепенные.
, что уменьшает значение биомассы (73, 74). По этим индексам виды делились на доминантные (руководящие), субдоминантные и второстепенные.
Л.А. Зенкевич и В.А. Броцкая, предлагая извлекать квадратный корень из величины
BP, имели ввиду только неудобство оперирования большими числами, особенно при построении графиков. Но эта операция имеет и более глубокий смысл. В рассматриваемом аспекте биомасса и встречаемость являются величинами не аддитивными, а мультипликативными, т.е. значимость вида пропорциональна произведению этих величин, а не их сумме. Именно для осреднения таких величин применяется средняя геометрическая, а не средняя арифметическая (5), т.е. индекс Броцкой - Зенкевича - это средняя геометрическая биомассы и встречаемости. Для оценки доминирования обе величины лучше брать в процентах. тогда индекс будет безразмерным с колебаниями от 0 до 100%. для оценки же детерминирования биомассу лучше брать в абсолютных единицах, поэтому индекс будет иметь определенную размерность, например, если биомасса берется в г/м2 , то индекс будет г0.5 м--1.В качестве индекса доминирования используется и произведение
BP без извлечения корня (33, 35, 89), а также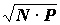 (53, 76, 87). Название этих индексов “коэффициентами репрезентативности” (76) неудачно, поскольку термин “ репрезентативность” имеет уже общепринятую статистическую трактовку. В.И. Лапшина применяет следующие градации степени доминирования (53):
(53, 76, 87). Название этих индексов “коэффициентами репрезентативности” (76) неудачно, поскольку термин “ репрезентативность” имеет уже общепринятую статистическую трактовку. В.И. Лапшина применяет следующие градации степени доминирования (53):

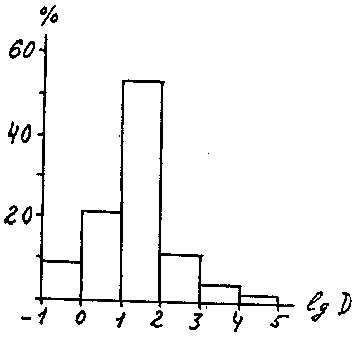
Рис. 8. Распределение видов по классам доминирования.
По оси абсцисс – классы доминирования, по оси ординат – процент видов, относящихся к данному классу
В последнее время в литературе по пресноводной гидробиологии встречается индекс доминирования, предложенным польским исследователем А. Ковнацким (141), в наших обозначениях он будет:
D
= 100 P · ni / S ni.Если принять во внимание, что
ni/S ni - это относительное обилие, то видно, что индекс Ковнацкого аналогичен “коэффициенту обилия” Палия (85), K = NP / 100, но в первом случае N и P выражают в долях единицы, во втором в процентах. А. Ковнацкий предложил следующую грацию доминирования по значению его индекса: 100 - 10 - доминанты, 10 - 1 - субдоминанты, 1 - 0.1 - адоминанты группы А, 0.1 - 0.01 - адоминанты группы В. Этот индекс применял В.А. Любин (60), по ошибке назвав его таксоценом; И.К. Тодераш по этому показателю выделял доминантные, субдоминантные и подчиненные формы с параллельным выделением руководящих, характерных и второстепенных видов по индексу Броцкой - Зенкевича (100).Рекомендуя применять этот индекс в экологических исследованиях, Б.Г. Иоганзен и Л.В. Файзова (40) не совсем удачно поместили его формулу в текст, так что создается впечатление, что формула предложена П.В. Тузовским.
Трехпараметрический индекс Л.В. Арнольди (2, 3) (
BNP)0.333 и “популяционный коэффициент” (40, 43)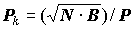 не получили широкого распространения.
В зависимости от применяемого показателя ранг доминирования вида в сообществе может сильно меняться, что видно из сравнения рис. 5 (доминирование по численности), рис. 9а (доминирование по биомассе) и рис. 9б (доминирование по индексу Броцкой - Зенкевича). Если в первом случае Paratanytarsus sp. занимает первое место, то во втором - шестое, в третьем - пятое. Лишь вид Stictochironomus crassiforceps везде занимает второе место.
не получили широкого распространения.
В зависимости от применяемого показателя ранг доминирования вида в сообществе может сильно меняться, что видно из сравнения рис. 5 (доминирование по численности), рис. 9а (доминирование по биомассе) и рис. 9б (доминирование по индексу Броцкой - Зенкевича). Если в первом случае Paratanytarsus sp. занимает первое место, то во втором - шестое, в третьем - пятое. Лишь вид Stictochironomus crassiforceps везде занимает второе место.
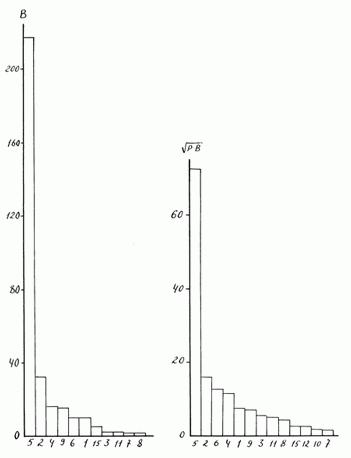
Рис. 9. Характер доминирования хирономид в оз. Плещеево
По оси абсцисс – номера видов, по оси ординат – индексы доминирования –
Т. Э. А. Фрей (110) предложил индекс
FCW, для нахождения которого пробы берутся большими квадратами. разделенными на ряд секций. Затем находят FCW = K·P¦ c·W, где P¦ c - вероятность нахождения вида в какой-либо секции квадрата, W - продуктивность вида (для растений он брал вес наземных частей) - константа. Этот индекс назван им “индексом постоянства доминантности. В дальнейшем было показано (20), что индекс Фрея фактически равен произведению биомассы на встречаемость.Большое количество разнообразных эмпирических индексов значимости применяется в фитоценологии. Дикстерхьюз (цит. по 12) использовал индекс П·
P, где П - проективное покрытие, его же использовали В.М. Понятовская и И.В. Сырокомская (89), а в дополнении к нему - индексы Р·В и П·В, причем они вычислялись как по абсолютным, так и по относительным значениям параметров.Используется также величина
N·P (14), индекс DFD = N + По + P, где N -относительная численность, По - относительная площадь сечения стволов данного вида, P - относительная встречаемость (126), в дальнейшем этот индекс получил обозначение IVI (127). Линдсей (цит. по 133) добавил к этому индексу четвертый компонент - относительное проектированное покрытие. Каяма ввел два индекса - B(N+P+П) / 3 и B(N+P+П+H) / 4, где Н - относительная высота растения (139). Эти индексы не нашли широкого применения в силу своей полной произвольности и отсутствия биологического содержания (70). Для глазомерной оценки значимости видов применяется бальная шкала “ДОМИН”, использующая показатели обилия, проективного покрытия и мощности развития растения (70).Один из последних индексов (75)
V
= 100g/(n(mz)0.5)где
g - сумма встречаемости видов данной группы, n - число описаний пробных площадок, m - среднее число видов на пробной площадке, z - число видов в данной группе. Этот индекс помогает оценивать значимость таксономической или экологической группы видов в составе растительного сообщества.Де Фриз ввел показатель, названный им “ порядком доминирования”,
Dt = 100 DF / PF где DF - частота доминирования, о которой говорилось выше, PF - частота встречаемости (12, 46, 61, 129, 140). При его методе пробы отбираются на равных расстояниях вдоль параллельных трансект, в зависимости от величины индекса проба обозначается определены значком: > - I место, < - II место, = - III место, : - прочие места. Б.А. Быков (16, 17) указал на важность учета устойчивости показателя доминирования во времени, он предложил индекс доминирования D = Gc · Ic / 100Ib, где Gc - участие вида в сообществе по биомассе, Ic - индекс устойчивости данного вида, Ib - индекс устойчивости всех видов. В качестве индекса устойчивости он считал возможным брать коэффициент вариации биомассы вида по многолетним данным.Неоднократно указывалось, что в показатель доминирования необходимо ввести параметр, характеризующий пространственное распределение организмов, одной из реализаций этого требования является учет встречаемости организмов в пробах, но встречаемость учитывает только факт наличия данного вида без характеристики его количественной представленности. Кроме того, чем больше встречаемость, тем больше получается величина индекса доминирования. оцениваемого. например, индексом Броцкой - Зенкевича. Это оправдано при оценке детерминирующего действия вида, поскольку, чем в большем количестве проб встречается вид при данной биомассе, тем большее влияние он оказывает на сообщество. Но в некоторых ситуациях подход должен быть иным. Например, при оценке степени доминирования кормовых организмов было показано, что увеличение степени неравномерности распределения пищи увеличивает ее доступность для потребителя. Увеличение агрегированности кормовых организмов эквивалентно увеличению их биомассы (38). Для оценки агрегированности предложено большое количество различных индексов (5, 93), наиболее удобным для нашей цели является индекс
A = х/хг, где х - среднее арифметическое, хг - среднее геометрическое, он обратен “показателю неоднородности структуры сообщества” (107). При равномерном распределении организмов его величина равна единице, при увеличении агрегированности - увеличивается. Индекс доминирования получает вид DА = A · B, он имеет размерность биомассы. Его стандартная ошибка SD = (SA2 + SB2)0.5 , стандартная ошибка биомассы определяется обычным методам статистики, а агрегированности - по формуле SA = (S2x + S2xг)0.5, метод оценки Sхг изложен в работе (5). К сожалению, вышеописанный индекс агрегированности не может быть вычислен при наличии в вариационном ряду нулевых значений. При большом количестве нулей их принято заменять единицами, но когда нулей много, оценка получается сильно искаженной, в этом случае в качестве индекса агрегированности можно рекомендовать “измеритель концентрации классификации” (166)l = å Pi2.
Для данной цели он применяется следующим образом. Вариационный ряд для каждого вида делится на 10 классов от наименьшего до наибольшего значения с равными интервалами, затем подсчитывается количество цифр, попавших в каждый класс и находится их доля от общего числа вариант - Pi. Затем находят индекс агрегированности AY = 10 å Pi2. Множитель 10 введен для удобства дальнейших расчетов. Величина индекса изменяется от 1 (равномерное распределение проб по классам) до 10 (все пробы относятся к одному классу). Стандартная ошибка индекса приблизительно равна (166) SA = 20{[(S pi3-(S pi2)2]/N}0.5, где N - количество проб в выборке. Тогда индекс доминирования имеет простой вид, DY = AY · B. Если биомассу выразить в относительных величинах - процентах от общей биомассы всех видов, то индекс будет безразмерной величиной, его стандартная ошибка (SAy2 + SB2). Сравнивая ранги доминирования видов по этому индексу с таковыми, найденные по индексу Бродской - Зенкевича (рис. 9 и рис. 10), видно что у некоторых видов ранги изменились, например, виды № 6 и № 9 поменялись местами. уменьшился разрыв между видами № 5 и № 2.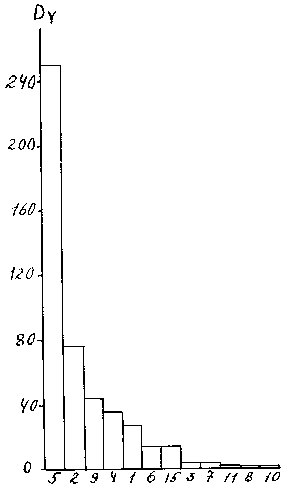
Рис. 10. Доминирование отдельных видов хирономид в оз. Плещеево, оцененное индексом
DYБ. Индексы структуры доминирования.
Используемые организмами ресурсы сообщества определенным образом делятся между слагающими его видами, что находит отражение в типах ранговых распределений видов. Эти распределения связывают показатели значимости (доминирования) видов с рангом - порядковым номером вида в общем ряду ранжированном по убыванию или возрастанию значимости. Предложено много математических моделей для описания ранговых распределений (56, 86, 109). Первой удачной моделью было логарифмическое распределение (131). К числу “классических распределений” относится также логнормальная модель (154), три распределения Макартура (145), называемые моделями “разломанного стержня”, геометрическое распределение (150). В последнее время предложен ряд новых моделей, таких как “обобщенный закон Ципфа - Мандельброта” (84), экспоненциальная модель (143), гиперболическая модель (54), модель “экспоненциально разломанного стержня”(108) дзета – модель (55). Эти модели исходят из определенных биологических предпосылок, поэтому обнаружение соответствия эмпирического материала математической модели позволяет сделать определенные выводы о процессах, протекающих в экосистеме, например, об обеспеченности организмов пищей, о наличии неблагоприятного воздействия загрязнителей и т. п. (109). Ранговое распределение численности отдельных видов хирономид в оз. Плещеево соответствует геометрическому распределению Мотомуры (рис. 11). Эта модель исходит из гипотезы “перехвата ниш”. Наиболее значимый вид занимает С - часть пространства ресурсов, второй вид занимает С - часть пространства ресурсов, оставшихся от первого вида, и т. д. Численность видов в этом случае представляет геометрическую прогрессию, в нашем примере численность последующего вида равна примерно 0.65 численности предыдущего. Это свидетельствует о наличии достаточно сильной конкуренции в сообществе.
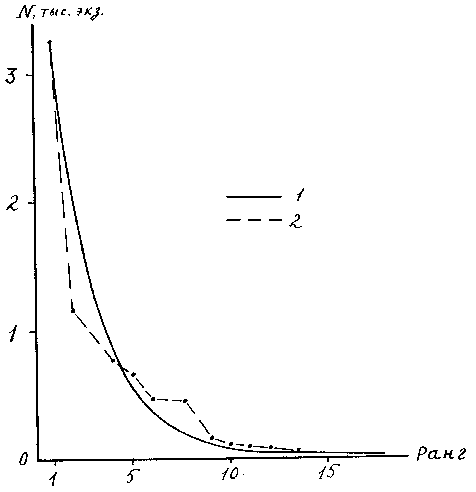
Рис. 11. Ранговое распределение численности хирономид в оз. Плещеево
1 – теоретическая кривая, 2 – эмпирические данные
Если эмпирическое распределение показателей значимости соответствует одному из теоретических распределений, то его параметры могут служить индексами, характеризующими структуру доминирования в сообществе. Пусть в двух выборках распределение численности видов соответствует модели Мотомуры, но в первой выборке С = 0.65, а во второй - 0.5, тогда можно сделать вывод, что во втором случае доминирование проявляется менее резко. Однако, проверка соответствия теоретическим распределениям достаточно сложна и требует сравнительно большого эмпирического материала. Малые же выборки, часто встречающиеся в полевой экологии, могут показать соответствие одновременно нескольким теоретическим моделям, основанным на совершенно разных предпосылках, поскольку при малой длине ряда разрешающая сила многих статистических критериев становится недостаточной. В таких случаях применяются более грубые методы описания характера доминирования в сообществе. Например, для этой цели может использоваться доля, которую оставляет первый по степени доминирования вид от суммарной численности (биомассы, продукции..) всех видов, т. е.
Dc = P1. Показано. что эта величина во многих случаях хорошо коррелирует с другими, более сложными показателями (124, 142, 146 ). Иногда в качестве индекса структуры доминирования берется отношение значимости первого вида ко второму, или же суммы первых двух видов к общему обилию (148, 153).Поскольку характер доминирования в сообществе определяется соотношением показателей доминирования отдельных видов (или иных групп), то в качестве индексов структуры доминирования могут выступать индексы агрегированности. Их разновидностью является широко используемые в экологии показатели “выровненности” или “эквитабельности”, обычно характеризующие степень равномерности распределения видов по обилию. Но в качестве исходных объектов могут быть взяты не только виды, которые могут характеризоваться и другими величинами, помимо обилия. Предложено большое количество таких показателей, среди исследователей нет единого мнения, какой из них предпочтительнее и в каких ситуациях.
Широко используется индекс
J = H/Hmax (153, 159), где H - информационный показатель Шеннона - Винера H = - S pi log pi . Критики этого индекса (122, 128, 142) отмечали, что его величина в сильной степени зависит от числа видов, при увеличении этого числа величина индекса стремится к единице, поэтому с его помощью нельзя сравнивать выборки, сильно различающиеся по числу видов. Далее, поскольку число обнаруженных видов всегда меньше, чем их имелось во всем сообществе, то оценка индекса получается смещенной, т. е. имеющей систематическую ошибку. Кроме того, при использовании этой формулы выборочное видовое разнообразие сравнивается с максимально возможным, а таковое возникает в ситуации, когда каждый вид представлен одинаковым количеством особей. Такая ситуация не реальна, поэтому было предложено (144) вместо теоретического максимального разнообразия использовать экологический максимум разнообразия , который получается, если распределение видов по обилию полностью соответствует модели № I “разломанного стержня” Макартура. Индекс выровненности при этом принимает вид E = S/S’, где S -фактическое число видов, S’ – число видов, которое наблюдалось бы при соответствии распределения вышеупомянутой модели Макартура, если бы при этом величина видового разнообразия оставалась без изменений.Более подходящими измерителями “выровненности” по мнению ряда исследователей может служить так называемое “модифицированное отношение Хилла” (122, 134 - 136); равное (
eн - 1)(S - 1), или показатель (H - Hmin)(Hmax - H) (128, 137). Поскольку при увеличении неоднородности показатели выровненности уменьшаются, то при использовании их как индексов структуры доминирования лучше брать обратные величины, т. е. D = 1/ E, где E -выровненность. Имеется строгое доказательство, что не существует такого индекса выровненности, который бы удовлетворял всем предъявленным к нему требованиям (157).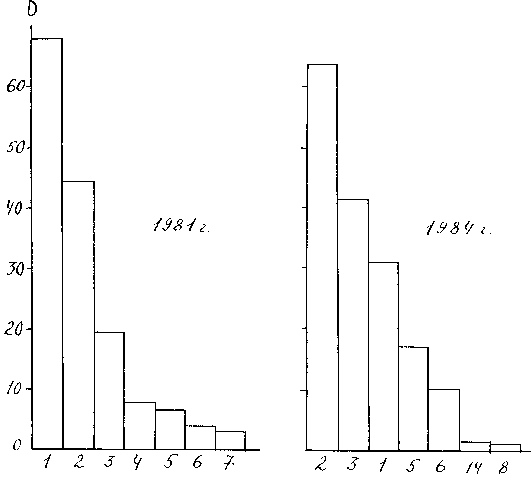
Рис. 12. Доминирование основных групп бентоса Чебоксарского водохранилища
, оцененное индексом Броцкой-ЗенкевичаРанее упоминавшийся “измеритель концентрации классификации” Юла широко используется в качестве индекса структуры доминирования и называется в этом случае индексом Симпсона (160, 163). Обычно его находят по формуле l
= S pi2, где Pi - доля данного вида в генеральной совокупности, ее несмещенная оценка для выборки будет l = [S ni(ni-1)]/[N(N-1)], где ni - число особей (или биомасса) i -го вида, N - число особей всех видов. Биологическая интерпретация этого индекса такова: l - это вероятность того, что две случайно отобранные особи будут относится к одному и тому же виду. При отсутствии доминирования величина индекса равна 1 / S, где S - число видов, при максимальном доминировании индекс равен единице. Для сравнения характера доминирования в двух выборках использованы данные по макрозообентосу Чебоксарского водохранилища, собранные осенью 19981 г. (первый год существования водохранилища) и 1984 г. (рис. 12 и 13). При этом рассматривалось не доминирование отдельных видов, а крупных таксономических групп, которых было выделено 14: 1. Личинки хирономид, 2. Олигохеты, 3. Кормовые моллюски, 4. Личинки коретры, 5. Гаммариды, 6. Пиявки, 7. Личинки комаров-мокрецов, 8. Водяные ослики, 9. Личинки поденок, 10. Личинки ручейников, 11. Личинки стрекоз, 12. Личинки прочих двукрылых, 13. Гусеницы бабочек, 14. Прочие.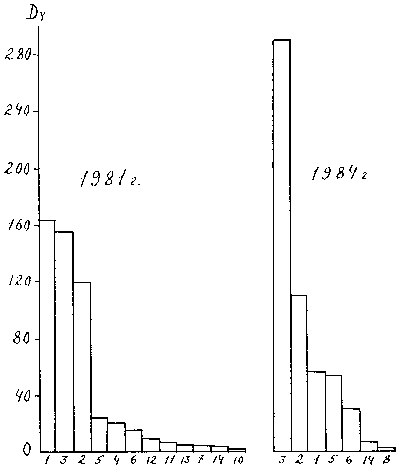
Рис. 13. Доминирование основных групп бентоса Чебоксарского водохранилища, оцененное индексом
DYНа рисунках предоставлены не все из этих групп, так как у некоторых их них индексы доминирования были столь малы, что изобразить их в принятом масштабе было невозможно. В качестве индекса индивидуального доминирования на рис. 12 использован индекс Броцкой-Зенкевича. причем биомасса бралась в процентах. на рис. 13 - наш индекс
DY = AY · B. Видно. что использование различных индексов существенно меняет представление о структуре доминирования. Из рис. 12 видно, что в 1984 г. по сравнению с 1981 г. индексы доминирования отдельных групп существенно изменились, на первое место вышли олигохеты, а хирономиды с первого места переместились на третье, но общая форма гистограммы изменилась незначительно. Для количественной оценки этого изменения в качестве индекса структуры доминирования мы использовали индекс Симпсона и коэффициент вариации индексов индивидуального доминирования отдельных групп. Оба показателя свидетельствуют о том, что разница между структурами доминирования в эти годы статистически недостоверна при уровне значимости 0.05. Гистограммы, представленные на рис. 13, визуально значительно отличаются друг от друга, но проверка по тем же показателям тоже показала недостоверность различий. В 1981 г. CV = 1.57 ± 0.3; l = 0.23 ± 0.23; а в 1984 г. - 2.02 ± 0.38 и 0.34 ± 0.052 соответственно. Для сравнения двух рядов показателей доминирования мы также использовали непараметрический ранговый критерий Уилкоксона (Т - критерий), применяемый для сравнения двух независимых выборок (98), который в другом руководстве (52) назван критерием Уайта. Он также показал недостоверность различий в структуре доминирования в эти годы.Индексы структуры доминирования абстрагируются от состава сообщества, т.е. они оценивают, в каком соотношении находятся различные виды или группы, не обращая внимание на то, какие это виды. Т. е. замена одного доминирующего вида другим с таким же показателем доминирования не меняет структуру доминирования. “Ведь храм остается храмом независимо от того, построен он из кирпича или камня” (62, стр. 207-208).
По структуре доминирования сообщества делятся на полидоминантные (доминируют сразу несколько видов или групп), к ним относятся , например, луговая растительность или тропические леса (70), и монодоминантные, в которых один вид резко выделяется по степени доминирования. Среди морских бентологов в этом случае более употребительны термины “плосковершинный биоценоз” и “островерхий биоценоз”(119), причем считается, что “островерхость” свидетельствует об однородности биотопа, а “плосковершинность” - о его гетерогенности. Шорыгинский “островерхий” биоценоз соответствует сообществу вида-эдификатора или “биогенному ценотипу” Линдрота, а “плосковершинный биоценоз “ - абиогенному или “плоскостному ценотипу” (81). Рассматривая ситуацию, изображенную на рис. 13, можно прийти к выводу , что в 1981 г. сообщество бентоса характеризовалось
“плосковершинностью”, а в 1984 г. - “островерхостью”, но учет ошибок репрезентативности и применение статистических критериев показывает поспешность такого суждения. Поэтому и к опубликованным в литературе данным об изменении характера доминирования тех или иных сообществ нужно относится с известной осторожностью. Во многих случаях наблюдаемые изменения могут быть статистически недостоверными, т. е. случайными.Для сравнения характера доминирования видов в двух сообществах может быть рекомендован также следующий прием (142, 152). Находится относительное (т.е. в процентах или долях от общего доминирования) доминирование первого вида , затем суммарное доминирование I и II видов и т.д. Получается семейство индексов I-D, II-D, III-D... Сообщество А имеет более высокий индекс структуры доминирования, чем сообщество Б, если для любого номера вида выполняется неравенство
i-Da ³ i-DБ.. Графически это изображается кривой накопленных показателей доминирования - кумулятой (рис. 14 ), для построения которой по оси абсцисс откладываются ранги видов в порядке убывания индексов доминирования, по оси ординат - накопленные величины доминирования соответствующего числа видов. Из рисунка видно , что кривая, характеризующая доминирование основных групп бентоса Чебоксарского водохранилища в 1984 г. располагается выше, чем кривая 1981 г. , т. е. концентрация доминирования в 1984 г. была выше, но кривые различаются незначительно. Можно полагать, что изображение доминирования таким способом более наглядно отражает его характер, чем традиционно применяемые гистограммы. Подобные кривые называют также” профилем внутреннего разнообразия” (152). Важно отметить, что в нашем примере эти кривые не пересекаются, хотя могут быть и ситуации с пересечением. Считается, что в случае пересечения сравнивать доминирование этим методом нельзя. Более того, даже сравнение разнообразия сообществ в этом случае с помощью индексов Шеннона - Винера, Симпсона, Макинтоша, Бриллюэна и т. п. также неправомерно, поскольку в этом случае упорядочивание сообществ по величине их разнообразия будет меняться в зависимости от выбранных индексов (142).Построение таких кривых отдельно для численности и биомассы бентоса помогает определить степень загрязнения отдельных биотопов. На среднезагрязненных участках кривые численности и биомассы очень близки, на слабозагрязненных - кривые биомассы располагаются выше кривых численности, на сильнозагрязненных – наоборот (162).
Для количественного сравнения подобных кривых можно воспользоваться индексом
Dc = 1- Ec, где Ес - индекс выровненности, рассчитываемой по формуле (125):Ec
= [ (d1 + 2d2 + 3d3 + ... +Sds) – 0.5] / 0.5S, где di - относительные индексы индивидуального доминирования отдельных видов, S - число видов. Данный индекс изменяется в пределах от 0 до 1. В нашем примере доминирования бентоса Чебоксарского водохранилища в 1981 г. характеризуется числом 0.68, а в 1984 г. - 0.78, т.е. увеличилось незначительно, на 14%.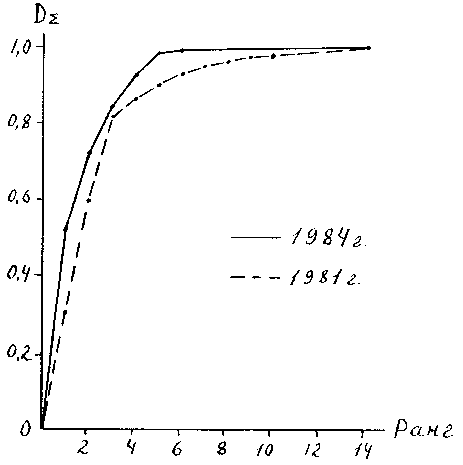
Рис. 14. Кумуляты доминирования бентоса Чебоксарского водохранилища
Для сравнения доминирования в нескольких сообществах может применятся показатель сходства, предложенный Ренконеном (151, 156). Численно он равен сумме минимальных индексов относительного доминирования двух выборок,
CD =S Dmin. Пусть имеются 3 выборки - А, B, С. Для каждого вида в каждой выборке найден какой-либо относительный (в процентах) показатель доминирования.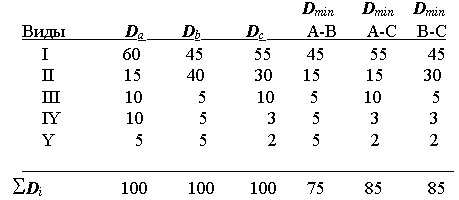
Следовательно, сходство по структуре доминирования между выборками А и В равно 75%, а выборка С имеет одинаковое сходство 85% с выборками А и В. Этот показатель использовал А. А. Шорыгин (118) для определения сходства пищевых рационов рыб, назвав его СП
- коэффициентом. Он считал. что СП - коэффициент характеризует “объем конкуренции” между изучаемыми видами рыб.III. СВЯЗЬ ДОМИНИРОВАНИЯ С ДРУГИМИ СТРУКТУРАМИ СООБЩЕСТВА.
Доминирование в той или иной степени связано с другими структурами, характеризующими сообщество. Например, временная структура проявляется и в изменении показателей доминирования. При сезонных изменениях биоценозов в силу особенностей биологии показатели обилия разных видов изменяются не синхронно, в результате чего происходит смена доминирующих видов (сменодоминантность). Гетеротопные организмы (проводящие разные периоды жизни в разных средах, например, личинки водных насекомых) и организмы, способные к миграциям, на определенный период могут исчезать из данного биотопа, так во многих
донных биоценозах пресноводных водоемов доминируют личинки хирономид, но летом в период массового вылета этих насекомых доминирование переходит к другим группам гидробионтов, обычно олигохетам. Изучая биоценозы бентоса Азовского моря, В. П. Воробьев (22) для отражения сезонной динамики доминирования ввел два показателя - коэффициент устойчивости биоценоза (процент станций, на которых выделенный по доминирующим видам биоценозов сохраняется от весны к осени, по отношению к числу станций, на которых он обнаружен весной) и коэффициент изменчивости биоценоза ( процент станций, на которых данный биоценоз от весны к осени переходит в другой биоценоз). Организмы, которые в момент изучения не доминируют, но способны стать доминантами в будущем, получили название футуруты (17).Особенно интересно прослеживать смену характера доминирования при экологических сукцессиях. При залитии водохранилищ в них образуются биоценозы, причем, если формирование донной фауны идет по I типу (4), то на первой стадии существования наблюдается резкое доминирование личинок мотыля. При изучении фитофильной фауны обнаружено, что в климаксных сообществах фитофильных беспозвоночных возрастает число видов-доминантов при уменьшении числа видов-сателлитов, а в “молодых” сообществах наоборот число видов-доминантов меньше, а сателлитов больше. Это влияет и на размерную структуру сообщества. У видов доминантов с увеличением размеров особей увеличивается вариабельность размеров, у видов-сателлитов - нет. При этом изменчивость размеров видов доминантов в климаксных сообществах значительно меньше, чем в “молодых” (37).
Смена доминирования при сукцессии тесно связана с жизненной стратегией видов, т.е. способами освоения территории, выживания и поддержания численности. Предложено несколько систем для выделения жизненных стратегий. Так, объединение систем Уиттекера и Раменского-Грайма позволяет различать пять стратегических типов (70): 1. Виоленты - виды, способные за счет активных биоморфологических реакций быстро захватывать и удерживать за собой пространство. 2. Экотопические патиенты - Виды, способные переносить абиотический стресс, обладающие развитым физиолого-биохимическим аппаратом оптимального использования скудных ресурсов. 3. Биоценотические патиенты - виды, испытывающие и приспособленные выдерживать биоценотический стресс со стороны конкурентов. 4. Ложные эксплеренты - виды, дающие вспышки численности во время кратковременного улучшения условий среды, например, однолетники пустынь, эфемероиды лесов. 5. Настоящие эксплеренты - виды, приспособленные жить в условиях нарушений среды, например, рудеральная растительность. Стратегия, оказавшаяся оптимальной относительно конкретных биотических и абиотических условий, позволяет виду получить высокий ранг доминирования. Настоящие эксплеренты в экологии называются также видами - оппортунистами. Они первыми занимают освободившееся место и обычно характеризуются быстрым размножением, ранним созреванием. высокой плодовитостью, простой структурой популяций. В ходе сукцессии доминирование постепенно переходит к более конкурентноспособным видам. В постоянно нестабильных биоценозах климакс не устанавливается и все время доминируют виды-оппортунисты. Это характерно, например, для осушной зоны водохранилищ.
В одних условиях могут доминировать виды-генералисты (эврибионты), адаптированные к большому числу размерностей многомерной экологической ниши, поэтому они реже сталкиваются с лимитирующей размерностью. В других условиях доминируют виды-специалисты (стенобионты), адаптированные к одной размерности, которая является лимитирующей в данный момент (149).
При изучении зоопланктона пресноводных водоемов Европейской части СССР выделено три группы доминантов (88). 1. Эвритопные формы с широким географическим распространением. 2. Узкоадаптированные к определенным экологическим условиям, но с широким географическим распространением. 3. С ограниченным биотипическим и географическим распространением.
Для глубоких водоемов более свойственна олигодоминантность, для мелководных - полидоминантность. В биоценозах видовое разнообразие и характер доминирования прямо связаны с эффективностью, с которой хищники предотвращают монополизацию основных ресурсов среды одним видом. Умеренное хищничество снижает плотность доминантов, увеличивая видовое разнообразие (81, 83). У хищных насекомых доминирование выражено слабое, чем у травоядных. Это можно трактовать таким образом, что популяции хищных насекомых в первую очередь лимитируются пищевыми ресурсами, а популяции травоядных - иными факторами (155).
Доминантов меньше там, где физические факторы среды экстремальны, например, северный лес может на 50% состоять из деревьев одного - двух- видов, в тропических же лесах доминировать могут более десятка видов (83). При наличии градиента факторов среды может наблюдаться и градиент показателей доминирования. Это показано на примере зоопланктонных биоценозов пресноводных водоемов, в которых наблюдается изменение показателя доминирования некоторых видов по мере продвижения с севера на юг (88). В общем ареале вида фитоценологи выделяют формационную область
- доминантную часть ареала, где вид создает формации (совокупности ассоциаций, возглавляемых одними и теми же доминантами), интеграционную область, где вид является субдоминантом, ассектаторную область, где вид входит в состав различных формаций в качестве ассектатора или инградиента, перфорационную область, где вид отсутствует (17).При изучении бентоса Азовского моря было обнаружено, что устойчивость биоценозов возрастает по мере увеличения их “островершинности”, т.е. увеличения “концентрации доминирования” (21). Чем выше биомасса организмов бентоса, тем проще и однообразнее его структура и тем большую роль играет один доминирующий вид. Когда биомасса этого вида составляет более 90% биомассы всего биоценоза, то этот биоценоз находится на пути превращения в моноценоз (79). Но изучив данные К.Н. Несиса (79), В.В. Черепанов (113) пришел к выводу, что для детритофагов нет связи между биомассой биоценоза и долей доминирующего вида, а для сестонофагов такая связь прослеживается. Но этот факт он трактует иным образом - не доля доминирующего вида увеличивается при увеличении общей биомассы биоценоза, а биомасса биоценоза возрастает при увеличении доли доминирующего вида. Такое объяснение восходит к “основному биоценотическому принципу” А. Тинеманна: чем однообразие условия существования и развития организмов в биоценозе, тем проще его структура и выше продуктивность. Связь характера доминирования со степенью однородности биотопа показана также на примере бентоса Баренцева моря (104).
В морских бентосных сообществах, в которых доминирующий вид оказывается и фонообразующим (образует дополнительные экологические ниши, субстрат, убежище и пищу для других организмов), число видов больше и биомасса выше, чем в биоценозах, где есть несколько доминирующих, но не фонообразующих видов (24). Аналитически показано существование зависимости между продуктивностью экосистемы и организованностью ее структуры. При этом максимум продуктивности достигается в случае абсолютного доминирования одного вида (90).
Много исследований посвящено изучению связи структуры доминирования и пищевой структуры морских донных биоценозов. Еще В.П. Соловьев отметил закономерное чередование представителей инфауны и эпифауны в распределении видов по их значимости в бентосе Азовского моря (22). В дальнейшем было принято более подробное деление организмов бентоса на пищевые группировки (94, 103): неподвижно прикрепленные сестонофаги жестких грунтов, неподвижно прикрепленные сестонофаги мягких (рыхлых) грунтов, подвижные сестонофаги мягких грунтов, собирающие
детрит виды, заглатывающие грунт виды, хищники, трупоеды, всеядные организмы. Было обнаружено, что при ранжировании видов по степени доминирования наблюдается чередование видов, относящихся к различным пищевым группировкам. Это объясняется тем, что пищевые группировки объединяют виды со сходной экологией питания, поэтому конкуренция между ними за пищевые ресурсы обострена. Поэтому внутри группировки может доминировать только один вид. Между представителями разных группировок конкуренция ослаблена, что и приводит к установлению в биоценозах наблюдаемого порядка доминирования, т.е. доминирующие виды подбираются по принципу минимизации конкуренции (104). Если же доминирующие виды относятся к одной пищевой группировке, то обычно к разным зоогеографическим группировкам (81). Разумеется, из этого правила наблюдаются исключения (78), а в некоторых случаях эта закономерность может считаться не биологической, а чисто статистической (80). Ее проявление тесно связано со степенью обеспеченности пищей донных беспозвоночных (9, 79, 80).Знание характера доминирования видов в различных абиотических и биотических условиях позволяет значительно глубже понять особенности биологии организмов. Второй аспект практической значимости работ по изучению доминирования заключается в том, что классификация сообществ организмов опирается на структуру доминирования. Особенно подробно методы выделения и классификации сообществ разработаны в фитоценологии, где применяются классификации фитоценозов, основанные на разных принципах: филогенетическая, физиономическая, флористическая, экологическая, индуктивная, дедуктивная, монотетическая, политетическая... В гидробиологии наиболее часто сообщества (называемые также биоценозами, комплексами, группировками...) выделяются по одному доминирующему виду, причем в качестве индексов доминирования обычно используют какой-либо вариант индекса Броцкой-Зенкевича. Для оценки степени сходства биоценозов используют различные коэффициенты сходства, иногда довольно экзотические, например, достоверность разницы между информационных индексами разнообразия по доминированию (27, 28). В таких случаях биоценоз называется по доминирующему организму, например, биоценоз дрейссены, биоценоз мотыля. В старых работах иногда биоценоз обозначался латинским названием вида (рода) с прибавлением окончания etum, например, биоценоз дрейссены - Dreissenetum (74). В работах морских бентосников для обозначения биоценоза использовалась дробь, в числителе которой помещалось название доминанта эпифауны, в знаменателе - инфауны (13), иногда биоценоз называется по первым двум доминирующим видам (35). Последовательно этого принципа придерживался и В.Д. Воробьев - даже если на двух близких станциях доминирования различные виды, он считал, что здесь два разных биоценоза. Но доминирование вида в конкретной пробе носит часто случайный характер, поэтому многие исследователи полагают, что помимо доминирующих видов необходимо учитывать и особенности биотопа. Л.В. Арнольди (2) название биоценоза представлял в виде дроби, в числителе которой были доминирующие виды, в знаменателе - краткая характеристика биотопа. Варианты биоценоза, встречающиеся на разных грунтах, К.Н. Несис объединял в единицы более низкого ранга - группировки, а биоценозы с доминированием разных видов одного рода - в единицы более высокого ранга - группы биоценозов (79), аналогичные “параллельным сообществам” Торсона (161). Внутрибиоценозные группировки В.П. Воробьев называл комплексами (22) и обозначал названием доминанта и субдоминанта (под термином “комплекс” разные гидробиологи понимали различные образования), этого же приннципа придерживались и другие исследователи (1, 23, 74). Д.А. Александров (1) предложил свою систему классификационных единиц (синтаксонов): ассоциация, формация, группа ассоциаций, класс ассоциаций, субассоциация, тип бентоса, которая пока не получила распространение.
Выделение сообществ по доминантам неоднократно подвергалось критике. В частности, указывалось, что доминанты часто имеют широкую экологическую амплитуду (эвритопны) поэтому выделенные по этому принципу сообщества могут значительно различаться по набору видов и по абиотическим характеристикам (1, 102). Роль доминантов подвержена значительным сезонным колебаниям (32, 70), поэтому приходится оговаривать, что классификация по доминантам пригодна для определенного момента времени, или же брать средние за длительный период показатели доминирования.
Более удовлетворительной считается использование на ряду с доминантами для классификации сообществ также и детерминантов или эдификаторов видов (1, 65, 82, 114). М.И. Киселева (44) считала, что доминирующий вид одновременно можно рассматривать и как вид-эдификатор, создающий своеобразные условия, характерные для данного биоценоза. В том случае когда плотность населения этого вида уменьшается, исчезают и специфические черты биоценоза, поэтому за центр биоценоза она принимала участок, на котором доминирующий вид имел наибольший индекс плотности, граница биоценоза проводилась там, где биомасса этого вида резко уменьшается, но индекс плотности все еще остается выше, чем у прочих видов.
Критикуя выделение сообществ животных на основе доминантов, Ю.И. Чернов (114) отмечал, что животные, как правило, являются слабыми эдификаторами, по набору видов-доминантов трудно судить о принципиальных особенностях структуры животного населения. Иногда однотипные сообщества могут не иметь общих видов, а разнотипные - давать высокую степень сходства. Широко известно явление экологического викариата. Он считал, что смена доминантных видов не может служить основанием для изменения статуса рассматриваемой группировки без решения вопроса о том, какие конкретные изменения в структуре животного населения и сообщества в целом вытекают из этой смены. “Можно сказать, что вид как таковой по отношению к структуре животного населения - случайный компонент” (115, стр. 435). “Не видимой состав, а преобладание и соотношение тех или иных жизненных форм характеризует сообщество” (42, стр. 122). Сходных взглядов придерживались и другие исследователи (10, 57).
Тип биоценоза часто выделяется по доминирующим экологическим группировкам, при этом за основу берутся пищевые группировки. Биоценозы с одинаковой пищевой структурой могут резко различаться по видовому составу (80). Различие этих структур хорошо подчеркивается теорией параллельных биоценозов (161): при движении с севера на юг в цепи параллельных биоценозов видовой состав закономерно меняется, трофическая структура остается более или менее постоянной. В.П. Турпаева (103) выделяла типы донных биоценозов и называла их по трофическим группировкам, например, “тип фильтраторов А”. Во многих случаях выделение биоценозов по доминирующему виду и по пищевой группировке может совпадать, поскольку часто доминирующими пищевыми группировкам являются те, к которым принадлежат доминирующие виды. При использовании этого принципа характеристика биоценоза выражается формулой, например, 84a + 6b + 4c + 0.4d +4e. Цифры показывают процент биомассы данной группировки от общей биомассы бентоса. 1,6% “прочих” организмов в формулу не входят. Обозначения пищевых группировок таковы (48): a - неподвижные сестонофаги, b - подвижные сестонофаги, c - собирающие детрит, d - грунтоядные, е - хищники. О.Г. Кусакин (50) выделял четыре литоральных сообществ следующим образом: I. Доминирует один вид. 2. Доминирует родственные виды. 3. Доминирует виды, принадлежащие к одной жизненной форме. 4. Доминируют виды, принадлежащие к одной трофической группировке.
Хотя в силу простоты принцип выделения сообществ по доминирующим видам или группам видов применяется наиболее часто, его можно рассматривать только как первое приближение. Для большей детализации необходимо использовать достаточно полный набор признаков, характеризующих сообщество и его биотоп, и полученные данные обрабатывать методами многомерной классификации. Необходимо иметь в виду, что распределение организмов и факторов среды, как правило, континуально, поэтому всякие границы условны, а выделение сообществ всегда будет иметь элемент субъективности, связанный с исходными концепциями, набором используемых признаков и методами обработки.
В заключении можно сказать, что изучение доминирования видов, таксонов, экологических групп, структуры доминирования сообществ необходимо как для дальнейшего развития экологической теории, так и для многих практических приложений. Эта проблема и в методологическом, и в методическом аспектах остается недостаточно разработанной и представляет широкое поле деятельности для исследователей разного профиля.
ЛИТЕРАТУРА
1. Александров Д.А. О структуре бентоса на мелководьях Белого моря в связи с проблемами его картирования // Пробл. охраны природы в бассейне Белого моря. Мурманск, 1984.
2. Арнольди Л.В. Материалы по количественному изучению зообентоса в Черном море // Тр. ЗИН АН СССР. 1941. т. 7, вып. 2.
3. Арнольди Л.В. Материалы по количественному изучению зообентоса Черного моря. II. Каркинитский залив // Тр. Севастопольский биол. станции. 1949. т. 7.
4. Баканов А.И. Количественное развитие зообентоса в водохранилищах Советского Союза. Борок, 1985. 84 с. Рукопись деп. в ВНИТИ, № 2968-85.
5. Баканов А.И., Сметанин М.М. Стандартные ошибки индексов агрегированности // Экология. 1980. № 2.
6. Беклемишев В.Н. Основные понятия биоценологии в приложении к животным компонентам наземных сообществ // Тр. по защите растений. 1931. т. 1, вып. 2.
7. Беклемишев В.Н. О классификации биоценологических (симфизиологических) связей // Бюл. МОИП, отдел. биол. 1951, т. 56, вып. 5.
8. Беклемишев В.Н. Термины и понятия, необходимые для количественного изучения популяций эктопаразитов и нидиколов // Зоол. журн. 1961. т. 40, № 2.
9. Беклемишев К.В., Возжинская В.Б., Нейман А.А. Обзор симпозиума “Биологическая структура морей и океанов. Биологические ресурсы морских водоемов и пути их использования // Биол. ресурсы водоемов, пути их реконструкции и использования, М. 1966.
10. Берман Д.И., Виленкин Б.Я. Некоторые принципы исследования сообществ // Количественные методы в экол. и биоценол. животных суши. Л. 1975.
11. Богоров В.Г. Исследование питания планктоядных рыб // Бюлл. ВНИРО. 19934, № 1.
12. Браун Д. Методы исследования и учета растительности. М. 1957.
13. Броцкая В.А. Зенкевич Л.А. Количественный учет донной фауны Баренцева моря // Тр. ВНИРО. 1939. т. 4.
14. Буторина Г.Н. Эколого-ценотический анализ кустарничко-травяного яруса лесных ассоциаций // Типы лесов Сибири. М. 1963.
15. Быков Б.А. Доминанты растительного покрова Советского Союза. Алма-Ата. т. I - 3. I960-I965.
I6. Быков Б. А. Введение в фитоценологию. Алма-Ата. I970.
I7. Быков Б. А. Экологический словарь. Алма-Ата. I983.
I8. Вайнштейн Б. А. О статистической достоверности количественных учетов пресноводных беспозвоночных // Биол. внутр. вод. Информ. бюлл. I969. № 3.
I9. Вайнштейн Б. А. Об оценке сходства между биоценозами // Биол., морфол. и систематика вод. организмов. Л. I976.
20. Василевич В. И. Статистические методы в геоботанике Л. I969.
21. Виленкин Б. Я. Структура простых донных сообществ // Донная фауна краевых морей СССР. М. I976.
22. Воробьев В.П. Бентос Азовского моря // Тр. АзЧерНИРО. 1949. вып. 13.
23. Гейнрих А.К. О приповерхостном планктоне северо-восточной части Тихого океана // Тр. ИО АН СССР. 1964. т. 65.
24. Голиков А.Н., Скарлато О.А. Некоторые принципы комплексного ландшафтно-географического районирования шельфа на экологической основе // Биол. моря. 1979. т. 5.
25. Грейт-Смит П. Количественная экология растений. М. 1967.
26. Дажо Р. Основы экологии. М. 1975.
27. Денисов Н.Е. Некоторые вопросы методики водолазных исследований донных сообществ // Океанология. 1972. т. 12, № 5.
28. Денисов Н.Е. Выделение и картографирование донных сообществ сублиторали по данным водолазных количественных исследований: Афтореф. дис. ...канд. биол. наук. М., 1974.
29. Житина Л.С. Пространственное распределение фитопланктона Белого моря: Автореф. дис. ...канд. биол. наук. М., 1981.
30. Зайцев Г.Н. Построение шкал балльной оценки // Биометрические методы. М. 1974.
31. Зайцев Г.Н. Сравнение флор по их видовому составу // Прикладная математика в биологии. М. 1979.
32. Зарубин С.Н. Определение доверительных значений коэффициентов сходства фитоценозов // Колич. методы анализа растительности. Т. 2. Рига. 1971.
33. Зацепин В.И., Зенкевич Л.А., Филатова З.А. Материалы по количественному учету донной фауны литорали Кольского залива // Тр. ГОИН. 1948. вып. 6 (18).
34. Зенкевич Л.А. Броцкая В.А. Материалы по экологии руководящих форм бентоса Баренцова моря // Уч. записки МГУ. Зоология. 1937. вып. 13, № 3.
35. Зимбалевская Л.Н. Сообщества беспозвоночных в зарослях высшей водной растительности среднего Днепра. // Гидробиол. журн. 1964. т. 1, № 3.
36. Зимбалевская Л.Н. Распределение фитофильных беспозвоночных и методы их количественного учета (Сообщение 2) // Гидробиол. журн. 1973. т.9, № 6.
37. Зимбалевская Л.Н. Изменчивость размеров пресноводных фитофильных беспозвоночных как проявление структуры их сообществ // Докл. АН СССР. 1975. т. 223, №
3.38. Ивлев В.С. Экспериментальная экология питания рыб. Киев. 1977.
39. Иоганзен Б.Г. Основы экологии. Томск. 1959.
40. Иоганзен Б.Г., Файзова Л.В. Об определении показателей встречаемости, обилия, биомассы и их соотношения у некоторых гидробионтов // Тр. ВГБО. 1978. т. 22.
41. Ипатов В.С., Кирикова Л.А., Линдеман Т.Н. Об оценке степени участия видов в структуре растительного покрова // Ботанич. журн. 1966. т. 51, № 8.
42. Кашкаров Д.Н. Среда и сообщество. М. 1933.
43. Кесминас В.А. Учет видового состава, численности и биомассы рыб в разных биотопах бассейна р. Мяркис // Матер. 14 (22) заседания советской рабочей группы по проекту № 8б “Вид и его продуктивность в ареале”. Вильнюс. 1984.
44. Киселева М.И. Структура биоценозов рыхлых грунтов Черного моря: Афтореф. дисс. ...канд. биол. наук. Севастополь, 1976.
45. Ковальчук А.А. О возможности использования численно- энергетического подхода при выращивании таксоценозов бентосных инфузорий // Биоценол. исслед. на Украине. Тезисы докл. - Львов. - 1984.
46. Кожова О.М. Формирование фитопланктона Братского // Формирование природ. условий и жизни Братского в-ща. М. 1970.
47. Константинов А.С. Общая гидробиология . М., 1979.
48. Кузнецов А.П. Экология донных сообществ мирового океана. М., 1980.
49. Куркин К.А. Системный подход в экологическом исследовании // Систем. исследования., 1977.
50. Кусакин О.Г. Литоральные сообщества // Океанология. Биология океана. т. 2. М., 1977.
51. Кучерук Н.В., Савилова Т.А. Количественная и экологическая характеристика донной фауны шельфа верхнего склона района северо-перуанского апвелинга // Бюлл. МОИП. Отдел биол. 1985. т.90, вып. 6.
52. Лакин Г.Ф. Биометрия. М., 1980.
53. Лапшина В.И. Использование модификации индекса плотности в исследованиях планктона // Изв. ТИРО. 1982. т. 106.
54. Левич А.П. Структура экологических сообществ //Биол. науки. 1977. № 10.
55. Левич А.Г. Ранговые распределения численности фитопланктона Белого моря // Биол. науки. 1979. № 4.
56. Левич А.п. Структура экологических сообществ. М., 1980.
57. Левушкин С.И. Понятие вида и экология // Матер. к II Всес. совещ. “Вид и его продуктивность в ареале”. Вильнюс, 1976.
58. Леме Ж. Основы биогеографии. М., 1976.
59. Любарский Е.Л. К методике экспресс-квалификации и сравнения описаний фитоценозов // Колич. методы анализа растительности. Уфа, 1974.
60. Любин В.А. Изменения в составе фауны малощетинковых червей Куйбышевского водохранилища // Гидробиол. журн. 1974. т. 10, № 6.
61. Макаревич В.Н. Голландские методы учета обилия видов по Де Фрису в сравнении с другими методами определения участия видов в луговых травостоях // Ботан. журн. 1966. т. 51, № 2.
62. Макфедьен Э. Экология животных. М., 1965.
63. Мина М.В. Рост животных // Зоол. позвоночных. т. 4. Итоги науки и техники ВНИТИ. М., 1973.
64. Мина М.В., Клевезаль Г.А. Рост животных. М., 1976.
65. Миркин Б.М. Критерии доминантов и детерминантов при классификации фитоценозов // Ботан. журн. 1968. т. 53, № 6.
66. Миркин Б.М. Рецензия на книгу “Разумовский С. М. Закономерности динамики биоценозов” // Журн. общей биол. 1983. т. 44, № 1.
67. Миркин Б.М. Рецензия на книгу “Динамика ценопопуляций растений” // Журн. общей биол. 1986. т. 47. № 5.
68. Миркин Б. М., Мухаметшина В.С., Муст Н.М., Наумова Л.Г. Опыт классификации растительных сообществ по критериям доминантов-детерминантов с использованием результатов корреляционного анализа // Колич. методы анализа растительности. т.2. Рига, 1971.
69. Миркин Б.М., Розенберг Г.С. Фитоценология. М.,1978.
70. Миркин Б.М., Розенберг, Г.С. Толковый словарь современной фитоценологии. М.,1
983.71. Мирошниченко М.П. Структура и продуктивность биоценозов бентоса Цимлянского водохранилища // Сб. науч. тр. ГосНИОРХ. 1982. № 184.
72. Митропольский В.И., Мордухай-Болтовской Ф.Д. Макробентос // Методика изуч. биогеоценозов внутр. водоемов. М., 197
5.73. Мордухай-Болтовской Ф.Д. Состав и распределение донной фауны в водоемах дельты Дона // Тр. АзЧерНИРО. 1940. Вып. 12. Ч. 2.
74. Мордухай-Болтовской Ф.Д. Материалы по гидробиологии Миусского лимана // Уч. зап. РГУ. 1948. т. 12.
75. Наткевичайте-Иванаускене М.П. Количественные соотношения встречаемости и константности групп видов в растительных сообществах // Биол. науки. 1985. № 8.
76. Наумов А.Г. Биогеографическая природа руководящих видов зоопланктона Тихоокеанского сектора Южного океана // Тр. ВНИРО. 1973. Т. 84, ВЫП. 4.
77. Наумов Н.П. Экология животных. М.. 1963.
78. Нейман А.А. О закономерностях состава морских донных биоценозов // Зоол. журн. 1963. т. 42. № 4.
79. Несис К.Н. Биоценозы и биомасса бентоса Нъюфаунлендско-Лабрадорского района // Тр. ВНИРО. 1965, Т. 57.
80. Несис К.Н. Некоторые вопросы пищевой структуры морского биоценоза // Океанология. т. 5, вып. 4.
81. Несис К.Н. Общие экологические понятия в приложении к морским сообществам. Сообщество как континуум. // Океанология. Биол. океана. т. 2. М., 1977.
82. Норин Б.Н. Совещание по методам выделения растительных ассоциаций 15-17 мая 1967 г. в Ленинграде // Ботан. журн. 1968. т. 53, № 6.
83. Одум Ю. Основы экологии. М., 1975.
84. Орлов Ю.К. Обобщенный закон Ципфа-Мандельброта и частотные структуры информационных единиц различных уровней // Вычислительная лингвистика. М., 1976.
85. Палий В.Ф. О количественных показателях при обработке фаунистических материалов // Зоол. журн. 1961. т. 40, вып 1.
86. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М., 1982.
87. Петров К.М. Биоценозы рыхлых грунтов черноморской части подводного склона Таманского полуострова // Зоол. журн. 1961. т. 40, вып. 3.
88. Пидгайко М.Л. Зоопланктон водоемов европейской части СССР. М., 1984.
89. Понятовская В.М., Сырокомская И.В. Опыт сравнительной оценки участия вида в строении лугового сообщества // Геоботаника. 1960. вып. 12.
90. Пузаченко Ю.Г. Соотношение между структурой и продуктивностью в биогеоценотических системах // Биол. системы в земледелии и лесоводстве. М., 1974.
91. Работнов Т.А. О структурных элементах фитоценозов и фитоценотических популяциях // Биол. МОИП .Отдел. биол. 1985. т. 90, вып. 1.
92. Реймерс Н.Ф., Яблоков А.В. Словарь терминов и понятий связанных с охраной живой природы. М., 1982.
93. Розенберг Г.С. Обзор методов статистической геоботаники. 6. Мозаичность. М., 1978. Рукопись деп. в ВНИТИ, № 46-78.
94. Савилов А.И. Экологическая характеристика донных сообществ беспозвоночных Охотского моря // Тр. ИО АН СССР.1961. т. 46.
95. Семкин Б.И. О постановке задач в структурной биоценологии //Теоретико-графовые методы биогеграф. исслед. Владивосток, 1983.
96. Семкин Б.И. Современная математика и фитоценология //Колич. методы анализа растительности. Уфа, 1974.
97
. Сметанин М.М., Баканов А.И. Об использовании средней арифметической и средней геометрической в гидробиологических и ихтиологических исследованиях // Биол. внутр. вод. Информ. бюл. 19980, № 48.98. Терентьев П.В., Ростова Н.С. Практикум по биометрии. Л.,
1977.99. Тимофеев-Ресовский Н.В. Выступление в дискуссии по теме “Определение понятия системы и системного подхода” // Системные исследования. М., 1970.
100. Тодераш И.К. Функциональное значение хирономид в экосистемах водоемов Молдавии. Кишинев, 1984.
101. Трасс Х.Х. О типологии доминантов растительных сообществ // Бюлл. МОИП. Отдел. биол. 1963. т. 48, вып. 15.
102. Трасс Х.Х. О значении доминантов растительных сообществ для классификации растительного покрова // Пробл. соврем. ботаники. т.1. М. Л., 196
5.103. Турпаева Е.П. Типы морских донных биоценозов и зависимость их распределения от абиотических факторов среды // Тр. ИО АН СССР. 1954. т. 2.
104. Турпаева Е.П. Пищевые взаимоотношения между доминирующими видами в морских донных биоценозах // Тр. ИО АН СССР. 1957. т. 20.
105. Уиттекер Р. Сообщества и экосистемы. М., 1980.
106. Федоров В.Д. Доминирующие формы фитопланктона Белого моря // Докл. АН СССР. 1969. Т. 188, № 4.
107. Федоров В.Д. Новый показатель неоднородности структуры сообщества // Вестн. МГУ. Биол. почвовед. 1973. № 2.
108. Федоров В.Д. Относительное обилие симпатрических видов и модель экспоненциально разломанного стержня (ЭРС) // Человек и биосфера. Вып. 2 М., 1978.
109. Федоров В.Д. Гильманов Т.Г. Экология. М., 1980.
110. Фрей Т.Э.-А. Некоторые аспекты фитоценологической значимости вида в растительном сообществе // Ботан. журн. 1966. т. 51, № 8.
111. Фрей. Т.Э-А. Биоценометрия - отрасль науки, нуждающаяся в определении //Колич. методы анализа растительности. Татру, 1969.
112. Фридерикс К. Экологические основы прикладной зоологии и энтомологии. М.-Л., 1932.
113. Черепанов В.В. Несколько замечаний к работе К.Н. Несиса “Некоторые вопросы пищевой структуры морских биоценозов” // Океанология. 1967. т. 7, вып. 3.
114. Чернов Ю.И. О некоторых индексах, используемых при анализе структуры животного населения суши // Зоол. журн. 1971. т. 50. вып. 7.
115. Чернов Ю.И. Понятие “животное население” и принципы геозоологических исследований // Журн. общей биол. 1971. т. 32, № 4.
116. Шварц С.С., Пястолова О.А., Добринская Л.А., Рункова Г.Г. Эффект группы в популяциях водных животных и химическая экология. М.,1976.
117. Шенников А.П. Введение в геоботанику. Л., 1964.
118. Шорыгин А.А. Питание и пищевые взаимоотношения рыб Каспийского моря. М., 1952.
119.
Шорыгин А.А. О биоценозах // Бюлл. МОИП. Отдел. биол. 1955. т. 60, вып. 6.120. Эдельгауз Г.Е. Достоверность статистических показателей. М., 1977.
121. Яхонтов В.В. Экология насекомых. М., 1969.
122. Alatalo R.V. Problems in measurement of eveness in ecolo
gy // Oikos. v. 37, № 2.123. Arrhenius O. Species and area
// J. Ecol. 1921. v. 9, № 1.124. Berger W.H., Parker P.L. Diversity of planktonic Foraminifera in deep sea sediments // Science. 1970. V. 168, № 3937.
125. Cieslak M. Propozycja okreslania ctruktury dominacji i roznorodnosci gatunkowrj zespolow // Wiad. ekol. 1980. T. 26, z. 2.
126. Curtis J.T. The palo verde forest type near Gonivaves, Haiti, and its relation to the surrounding vegetation. // Caribbean For. 1947, V. 8.
127. Curtis J.T., McIntosh
R.P. An upland forest continuum in the prairie-forest region of Wisconsin // Ecology. 1951. V.32, № 2.128. De Benedictis P.A. On the correlations between certain diversity indices // Amer. Natur. 1973. V.107, № 954.
129. De Vries M. Methods used in plant sociology and agricultural botanical Grassland research // Herbage Rev. 1937. V. 5.
130. Fager E.W. Determination and analysis of recurrent groups // Ecology. 1957, V. 38, № 4.
131. Fisher R.A., Corbet A.S., Williams C.B. The relation between the number of species and the number of indi
viduals in a random sample of an animal population // J. animal ecol. 1943, V. 12, № 1.132. Gleason H.A. On the relation between species and area // Ecology. 1922. V.3, № 1.
133. Goff P.C., Cottam C. Gradient analysis: the use of species and synthetic indi
ces // Ecology. V. 48, № 5.134. Heip C. A new index measuring eveness // J. Mar. Biol. Assoc. U.K. 1974. V.54, № 3.
135. Heip C., Engels P. Comparing species diversity and eveness indices // J. Mar. Biol. Assoc. U.K. 1974.
V. 54, № 3.136. Hill M. C. Diversity and eveness: a unifying notation and its consequences // Ecology. 1973. V. 54, № 2.
137. Hurlbert S.N. The nonconcert of species diversity: a critique and alternative parameters // Ecology. 1971. V. 52, № 4.
138. Kasp
rzak K. Zgrupowania malzow z rodzaju Pisidium sensu lato (Bivalvia) w roznych typach zbiornikow wodnych // Fragm. faun. 1975. V. 20, № 9.139. Kayama R. New methods of quantitative representation of the structure of plant communities . IV. On the summed do
minance ratio weighted by the plant weight // Jap. J. Ecol. 1961. V.11, № 4.140. Kovachev S. C. , Usunov Y.J. Formation of macroinvertebrate communities in the course of the biological selfpurification of the Mesta River // Arch. Hydrobiol. 1986. Suppl. 72, H. 4.
141. Kownacki A. Taxocens of Chironomidae in streams of the Polish High Tatra Mts // Acta hydrobiol. 1971. V. 13, № 4.
142. Lambshead P. J. D., Platt H. M. The detection on of differences among assessment of marine benthic species based on an a
ssessment of dominance and diversity // J. Natur. Hist. 1983. V. 17, № 6.143. Lamont B.B., Downes S., Fox J.E.D. Importance-value curves and diversity indicts applied to a species-rich healthland in Western Australia // Nature. 1977. V. 265, № 5593.
144.
Lloyd M., Ghelardi R.J. A table for calculating the “equitability” component of species diversity // J. Animal. Ecol. 1964. V. 33, № 2.145. MacArthur R. H. On the relative abundance of bird species // Proc. Nat. Acad. Sci. USA. 1957. V. 43, № 3.
146. Мay
R. M. Patterns of species abundance and diversity // Ecol. and Evolution of Communities. Cambridge, 1975.147. McCloskey L.R. The dynamics of the community associated with a marine scleractinian coral // Intern. Rev. ges. Hydrobiol. 1970. Bd 55, H. I.
148.
McNaughton S.J. Relationships among functional properties of Californian grassland // Nature. 1967. V. 216, № 5111.149. McNaughton S. J., Wolf L. L. Dominance and the niche ecological systems // Science. 1970. V. 167, № 3915.
150. Motomura J. Etude statisique des populations ecologiques // Doobutugaku zassi (Tokyo). 1932. V.44.
151. Muller G. Bodenbiologie. Jena, 1965.
152. Patil G.P., Taillie C. Diversity as a concept and its implications for random communities // Bull. Intern. Stat. Inst. 1977. V. 47.
153. Pielou E. C. Mathematical ecology. N. Y. e. a., 1977.
154. Preston F.W. The commonnes and rarity of species // Ecology. 1948. V.29, № 3.
155. Pulliam H.R., Odum E.P., Barrett G.W. Equitability and resource limitation // Ecology. 1968. V.49, № 4.
156. Renkonen O. Statistisch-okologische Untersuchungen über die terrestricche Käferwelt der finnischen Bruchmoore // Ann. Zool. Soc. Zool. Bot. Fenn. Vanamo. 1938. V. 6, N 1.
157. Routledge R. D. Evenness indices: are any admissible? // Oikos. 1983. V.40, № 1.
158. Sanders H.L. Benthic studies in Buzzards Bay. III. T
he soft-bottom community. //Limnol. Oceanogr. 1960. V. 5. № 2.159. Sheldon A. L. Equitability indices: dependence on the species count // Ecol
ogy. 1969. V. 50, № 3.160. Simpson E.H. Measurement of diversity // Nature. 1949. V. 163, № 4148.
161. Thorson
G. Bottom communities (sublittoral or shallow shelf) // Met. geol. Soc. Amer. 1957. V.67, № 1.162. Warwick R.M. A new method for detecting pollution effects on marine macrobenthic communities // Mar. Biol. 1986. V. 92, № 4.
163. Whittaker R. H. Dominance and diversity in land plant communities // Sci
ence. 1965. V. 147, № 3655.164. Williams C. B. Patterns in the balance of nature and related problems in quantitative ecology. N. Y., 1964.
165. Yodzis P. The compartmentation of real and assembled ecosyste
ms // Amer. Natur. 1982, V. 120, № 5.166. Yule G. U. The statistical study of literary vocabulary. L., 1944.