© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Опубликовано: Актуальные проблемы герпетологии и токсинологии: Сборник научных трудов. Вып. 7. – Тольятти, 2004. – С. 45–59.
Cтатистический анализ флуктуирующей асимметрии билатеральных признаков разноцветной ящурки Eremias arguta
Д.Б. Гелашвили1, В.Н. Якимов1, В.В. Логинов1, Г.В. Епланова2
1Нижегородский государственный университет им. Н.И. Лобачевского, ecology@bio.unn.ru
2 Институт экологии Волжского бассейна РАН, г. Тольятти, ecology@avtograd.ru
Проблема симметрии (асимметрии) биологических объектов является одной из фундаментальных в современной биологии. Симметричным называется объект, который состоит из частей равных относительно какого-либо признака. Проверить равенство объектов или частей системы можно с помощью некоторого преобразования, которое совмещает равные объекты или части одного и того же объекта. Это означает, что объекты либо инвариантны относительно некоторых преобразований пространства, либо нет. Однако, в природе чаще всего встречаются лишь приблизительно симметричные объекты, об инвариантности которых относительно операций симметрии также можно говорить лишь приблизительно.
Симметрия, точная или приблизительная, является важнейшим свойством подавляющего числа живых организмов (Гиляров, 1944, Вейль, 1968; Урманцев, 1974; Захаров, 1987; Лима-де-Фариа, 1990; Аветисов, Гольданский, 1996; Марченко, 1999 и др.). При этом следует учитывать, что изменения структур и функций сравнительно независимы, т.е. морфофункциональная организация не жесткая система; конструкция имеет некоторый люфт в отношении каждой функции, и, наоборот, условия функционирования, задаваемые естественным отбором, допускают определенного масштаба селективно-нейтральные изменения структур (Гродницкий, 1998).
К такому типу изменений можно отнести флуктуирующую асимметрию (ФА), под которой понимают незначительные и случайные (ненаправленные) отклонения от строгой билатеральной симметрии биообъектов (Захаров, 1987). Таким образом ФА организмов по билатеральным признакам можно рассматривать как случайное макроскопическое событие, заключающееся в независимом проявлении либо на левой, либо на правой, либо на обеих сторонах тела, но в разной степени выраженных признаков, являющихся итогом стохастических микроскопических процессов. На макроскопическом уровне флуктуирующую асимметрию предлагают использовать в качестве меры в оценке стабильности развития организма (Захаров, 2001; Захаров и др., 2001). Уровень морфогенетических отклонений (т.е. ФА) от нормы оказывается минимальным лишь при определенных (оптимальных) условиях среды и неспецифически возрастает при любых стрессовых воздействиях. В качестве показателей стабильности развития обычно рассматривают нарушения развития (фенодевиации) и онтогенетический шум (Захаров и др., 2001).
Перспективы применения ФА в системе биомониторинга требует разработки корректных методов количественной оценки ее величины. Подходы к статистическому анализу ФА билатеральных признаков обобщены в работах А.В.Кожары (1985), В.М.Захарова (1987), В.М.Захарова и др. (2000), Д.Б.Гелашвили и др. (2001), Palmer, Strobeck (2003).
Число публикаций в мировой литературе, посвященных флуктуирующей асимметрии возрастает лавинообразно и сопровождается оживленной полемикой по методологическим проблемам и методическим аспектам стабильности (нестабильности) развития (Гелашвили, 2002; “Стабильность…”, 2002; Kozlov, 2003; Martel, Lempa, 2003; Palmer, Strobeck, 2003 и др.).
Обобщая сложившуюся в настоящее время ситуацию можно выделить три ключевых направления в исследовании ФА.
Первый из вышеперечисленных пунктов особенно важен при изучении и характеристике ФА билатеральных признаков у организмов, которые подвергаются анализу впервые. В данной работе будет рассмотрена ФА разноцветной ящурки Eremias arguta, самая северная популяция которой известна в окрестностях г. Тольятти. Этот аспект особенно интересен в связи с работой Ждановой (2003), показавшей, что у прыткой ящерицы Lacerta agilis уровень случайной изменчивости (ФА) возрастает в северной части ареала.
У разноцветной ящурки E. arguta статистическому анализу были подвергнуты два билатеральных признака: количество бедренных пор – P.f. и количество верхнегубных щитков – Lab. Животные были отловлены в 2001-2002 гг. Общий объем выборки составил 105 особей, из которых 63 – самцы и 42 – самки. Первичные данные, подвергнутые статистическому анализу, представлены в таблицах 1 (самцы) и 2 (самки).
В этих таблицах и далее в тексте приняты следующие обозначения и сокращения:
L и R, соответственно, левая и правая стороны тела; (L-R) и |L-R|, соответственно, величина асимметрии, то есть разность между величиной признака на левой и правой стороне тела с учетом знака и по абсолютной величине (по модулю); (L+R) и (L+R)/2, соответственно, суммарная и средняя величина признака на обеих сторонах тела. Все столбцы в таблицах 1 и 2 пронумерованы и на них даются ссылки при статистическом анализе первичных данных тем или иным методом.
Расчеты проведены с применением пакета "Statistica" и рекомендаций изложенных в работах Гланца (1999), Боровикова (2001а, 2001в), Ребровой (2002).
Уже предварительный анализ данных, приведенных в колонках 10 и 11 табл. 1 и 2, показывает довольно высокую частоту встречаемости асимметричных признаков в "тольяттинской" популяции E. arguta (табл. 3).
Таблица 3
Частота встречаемости асимметричных признаков в “тольяттинской” популяции разноцветной ящурки Eremias arguta

Однако этот результат ничего не говорит о типе асимметрии: флуктуирующей, направленной или антисимметрии. С канонических позиций математической статистики, ФА – это форма изменчивости различий между правой и левой сторонами, когда их значения (L-R) нормально распределены вокруг нуля. Направленная асимметрия отличается от флуктуирующей тем, что значение признака на одной из сторон в среднем больше, чем на другой. Статистически это выражается в отличии от нуля среднего различия между сторонами (L-R). Антисимметрия имеет место, когда асимметрия проявления признака является нормой, при этом совершенно не важно, в какую сторону направлено различие между сторонами. Статистически это выражается в том, что различия (L-R) распределены вокруг нуля, но частоты распределения отклоняются от нормальности в сторону отрицательного эксцесса или бимодальности (Захаров, 1987; Palmer, Strobeck, 1986; Palmer, Strobeck, 2003).
Из трех типов асимметрии адекватным индикатором нестабильности развития может служить только ФА (хотя дискуссии на эту тему продолжаются до сих пор, см. напр. Graham et. al., 1998). Поэтому прежде чем приступить к количественной оценке нестабильности развития через ФА необходимо убедится во флуктуирующем характере асимметрии каждого признака из выбранного набора. Эта процедура достаточно сложна,
требует применения различных методов статистического анализа. В статистическом анализе ФА билатеральных признаков можно выделить два аспекта. Первый из них связан с анализом индивидуальных признаков и включает:
Второй аспект связан с выбором и корректным применением интегральных показателей, оценивающих ФА выборки организмов по комплексу индивидуальных признаков в задачах биомониторинга.
Известно, что первым этапом анализа количественных данных является анализ вида их распределения. Реально в экспериментальных выборках, как правило, не бывает строго нормального распределения признака. Однако необходимо установить отобрана ли эта выборка из генеральной совокупности, в которой изучаемый признак имеет нормальное распределение. Решение этого вопроса определяет дальнейшую тактику статистического анализа: применение параметрических или непараметрических методов. Как указывает Гланц (1999, стр. 326):"… основная трудность состоит не в том, какой из перечисленных способов выбрать, а в том, что объем выборки слишком мал, чтобы применить любой из них. Убедительные свидетельства в пользу гипотезы нормальности или против нее встречаются редко. Гораздо чаще все решает интуиция, привычка и вкус исследователя". Не менее осторожны в оценке универсальности закона нормального распределения и такие выдающиеся математики как А. Пуанкаре и Р. Фишер. Так, А. Пуанкаре принадлежит высказывание: "Каждый уверен в справедливости нормального закона: экспериментаторы – потому, что они думают, что это математическая теорема; математики – потому, что они думают, что это экспериментальный факт" (цит. по Айвазян и др., 1983, С. 170). В свою очередь, Р. Фишер замечает: "Отклонения от нормального вида, если только они не слишком заметны, можно обнаружить лишь для больших выборок, однако сами по себе эти отклонения вносят малое отличие в статистические критерии и другие вопросы" (цит. по Боровикову, 2001, С. 131).
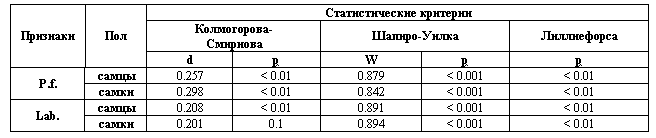
Тем не менее, в настоящее время существуют численные методы оценки соответствия изучаемого распределения признака нормальному, в том числе реализованные в пакете "Statistica" (критерии Колмогорова-Смирнова, Шапиро-Уилка, Лиллиефорса), позволяющие с достаточной строгостью обосновать выбор параметрических или непараметрических методов анализа.
Статистический анализ нормальности распределения значений асимметрии билатеральных признаков E. аrguta у самцов и самок (столбцы 6 и 7, табл. 1,2) показывает (табл. 4), что только признак Lab. у самок, судя по критерию Колмогорова-Смирнова, имеет нормальное распределение: р = 0.1; следовательно, нулевая гипотеза об отсутствии отличий от нормальности принимаются.
Таблица 4
Статистический анализ нормальности распределения значений асимметрии со знаком билатеральных признаков разноцветной ящурки Eremias arguta
Таким образом, применение непараметрических методов статистического анализа представляется вполне обоснованным. Среди таких методов (критериев), представленных в пакете "Statistica", для целей настоящей статьи применимы критерии Уилкоксона, Манна-Уитни, Крускала-Уоллиса. Критерий Уилкоксона применим для анализа данных связанных групп, т.е. полученных от одного объекта исследования (до и после лечения, справа и слева и т.п.) или от разных объектов, но подобранных в пары по определенным характеристикам. При сравнении двух групп по одному или нескольким количественным признакам, целесообразно применять критерий Манна-Уитни, а при числе групп три и более – Крускала-Уоллеса.
При проведении статистического анализа будем придерживаться вышеприведенной последовательности изучения параметров асимметрии индивидуальных признаков.
Для изучения направленности асимметрии признаков проведено сравнение данных представленных в столбцах 2-5 табл. 1 и 2 с помощью критерия Уилкоксона (табл. 5).
Таблица 5
Статистический анализ направленности асимметрии билатеральных признаков разноцветной ящурки Eremias arguta (критерий Уилкоксона)

Результаты анализа показывают, что ни в одном из случаев не установлено статистически значимых (р<0.05) различий в величине признака на левой и правой стороне тела. Другими словами, имеют место флуктуации асимметрии признака вокруг нулевого среднего, что является диагностическим признаком ФА.
Следующий шаг анализа связан с изучением зависимости величины асимметрии признака от его размера. При наличии этой зависимости с увеличением размера объекта может увеличиваться и величина асимметрии. Это можно объяснить тем, что в онтогенезе структуры, имеющие бóльшие линейные размеры, могут проявить и более выраженную асимметрию. Сильная размер-зависимость оказывает влияние как на форму распределения значений асимметрии, так и на интегральные характеристики выборки.
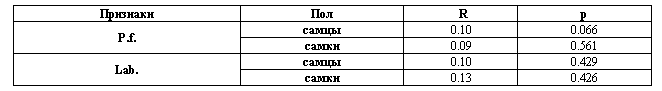
Для выявления размер-зависимости можно использовать непараметрический коэффициент ранговой корреляции Спирмена. В этом случае проверяется связь между абсолютными значениями асимметрии |L-R| и средним размером признака (L+R)/2, для чего используются данные, приведенные в столбцах 6-9 табл. 1 и 2. Обязательное использование непараметрических методов в данном случае обусловлено тем, что один из рядов, связь между которыми проверяется, а именно ряд абсолютных значений асимметрии, заведомо отклоняется от нормальности: он имеет так называемое усеченное нормальное или полунормальное распределение (Whitlock, 1996). Если положительная связь присутствует, в дальнейшем анализе нужно использовать прямое нормирование асимметрии на размер признака, либо использовать подразумевающие такое нормирование алгоритмы.
Результаты корреляционного анализа, приведенные в табл. 6, показывают отсутствие размер-зависимости у анализируемых признаков. Таким образом, в последующем анализе можно воспользоваться абсолютным значением асимметрии признака |L-R|. Однако для сопоставления результатов с данными, полученными другими исследователями, целесообразно ввести ту или иную нормировку. При этом следует учесть, что только нормировка вида |L-R|/(L+R) в отличие от |L-R|/[(L+R)/2], позволяет получить значение асимметрии в интервале 0÷1, удобном для сравнительного анализа.
Следующий шаг анализа позволяет обосновать включение изучаемого признака в систему признаков, интегрально оценивающих ФА организма. Очевидно, что чем менее скоррелированы величины асимметрии признаков, тем более независимую и объективную оценку можно получить с помощью данного набора признаков.
Таблица 6
Статистический анализ корреляции между абсолютными значениями асимметрии и средним размером признака разноцветной ящурки Eremias arguta (ранговый коэффициент корреляции Спирмена, R)
С этой целью проводится корреляционный анализ с применением рангового коэффициента Спирмена данных, приведенных в столбцах 2-7 табл. 1 и 2. Как и следовало ожидать, натуральные значения признаков на левой и правой сторонах тела тесно скоррелированы (см. строки 1 и 5 в табл. 7). В то же время натуральные значения разных признаков, равно как и величины их асимметрии, являются нескоррелированными между собой (табл. 7). Полученные результаты позволяют включить оба признака в систему интегральной оценки ФА выборки E. аrguta.
Таблица 7
Статистический анализ корреляции билатеральных признаков между собой и их асимметрии в популяциях разноцветной ящурки Eremias arguta (ранговый коэффициент корреляции Спирмена, R)

Важное значение имеет вопрос о наличии половых (гендерных) различий в средней величине асимметричных признаков, поскольку в случае их отсутствия можно использовать в анализе объединенную выборку самцов и самок. Применение критерия Манна-Уитни показало (табл. 8), что для обоих признаков гендерные различия отсутствуют. Таким образом, применение объединенных выборок для интегральной оценки ФА выборки E. аrguta обосновано.
Таблица 8
Статистический анализ гендерных различий асимметрии билатеральных признаков разноцветной ящурки Eremias arguta (критерий Манна-Уитни)

На последнем этапе проводятся так называемые тесты на идеальную ФА, позволяющие отличить ее от антисимметрии. Явление антисимметрии, также как и направленность, детерминируется генотипически. Антисимметрия не запрещает проявления ФА, однако получающуюся в результате сложную смесь двух типов асимметрии чрезвычайно сложно разделить. Поэтому от использования признака, проявляющего достоверную антисимметрию, лучше отказаться.
Поскольку антисимметрия проявляется в виде отрицательного эксцесса распределения различий между сторонами (L-R) либо (L-R)/(L+R)/2, то тестом на идеальную ФА является тест на значимость эксцесса. К сожалению, этот тест обладает серьезными недостатками: чувствительностью к малому размеру выборки и низкой мощностью. Поэтому можно воспользоваться табулированными критическими значениями эксцесса (Palmer, Strobeck, 2003) (табл. 9). Следует также отметить, что индикатором антисимметрии служит только отрицательный эксцесс, в то же время как положительный эксцесс указывает на генотипическую гетерогенность организмов выборки по стабильности развития (Whitlock, 1996; Whitlock, 1998; Van Dongen, 1998) и является нормой.
Таблица 9
Статистический анализ эмпирических (k) и критических (kcrit) значений эксцесса билатеральных признаков разноцветной ящурки Eremias arguta

Поскольку ни для одного из признаков, как у самцов, так и у самок, не выявлено превышения эмпирического значения эксцесса (k) над критическим (kcrit), можно считать, что антисимметрия у анализируемых признаков отсутствует.
После завершения анализа индивидуальных признаков можно переходить к интегральной оценке исследуемых выборок E. arguta по комплексу признаков. Поскольку в нашем распоряжении была выборка только из одного биотопа, сравнение проведем между выборками самцов и самок.
Использование в исследованиях флуктуирующей асимметрии только одного признака не позволяет делать надежные выводы, поэтому предпочтительно использование множества признаков. При этом каждый дополнительный признак добавляет одну степень свободы к оценке уровня нестабильности развития (Palmer, Strobeck, 2003). Следует отметить, что при использовании множества признаков, условия описанных выше этапов должны выполняться для всех из них.
В настоящее время известно большое число интегральных индексов ФА (см. обзор Palmer, Strobeck, 2003), из которых рассмотрим несколько, различающихся алгоритмами нормировки.
Введем следующие обозначения: Lij, Rij – значения j-го признака (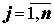 ) соответственно, слева и справа у i-й особи (
) соответственно, слева и справа у i-й особи (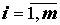 ), FAij – значение асимметрии j-го признака у i-й особи, bij – дискретизованное значение асимметрии (0 – отсутствие асимметрии j-го признака у i-й особи, 1 – наличие). Большая часть предложенных схем анализа флуктуирующей асимметрии множества признаков представляет собой комбинацию нескольких элементов, среди которых можно выделить нормирование данных, свертку информации и применение того или иного статистического критерия.
), FAij – значение асимметрии j-го признака у i-й особи, bij – дискретизованное значение асимметрии (0 – отсутствие асимметрии j-го признака у i-й особи, 1 – наличие). Большая часть предложенных схем анализа флуктуирующей асимметрии множества признаков представляет собой комбинацию нескольких элементов, среди которых можно выделить нормирование данных, свертку информации и применение того или иного статистического критерия.
Нормирование данных необходимо, когда выявлена значимая размер-зависимость асимметрии, либо когда разные признаки имеют разную размерность, а также когда имеет место значительная гетерогенность асимметрии признаков. Чаще всего нормировка производится следующим образом (Захаров и др., 2000):
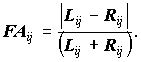
Принципиально иной метод нормировки:
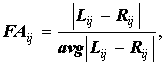
где avg|Lij-Rij| означает усреднение по всем выборкам, рассматриваемым в исследовании. Эта нормировка была предложена Leung et al. (2000), она направлена на то, чтобы снять гетерогенность асимметрии различных признаков. В результате применения такой нормировки значения асимметрии оказываются распределены вокруг единицы.
Наиболее простым и распространенным способом свертки является простое суммирование значений асимметрии всех признаков:
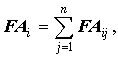
где n – число признаков.
В качестве альтернативы этому способу можно рассматривать недавно предложенный метод (Гелашвили и др., 2001), заимствованный из методов современной кристаллографии:
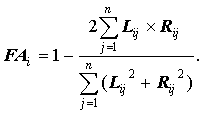
Особенностью этого метода является то, что он носит нелинейный характер, при этом нормировка производится одновременно со сверткой. Следует отметить, что этот метод нельзя применять в комбинации с нормировкой второго типа.
В таблице 10 приведены оценки уровня ФА выборок самцов и самок E. arguta (а также объединенной выборки), рассчитанные по алгоритмам, приведенных в работах разных авторов.
Таблица 10
Интегральные оценки флуктуирующей асимметрии выборок разноцветной ящурки Eremias arguta
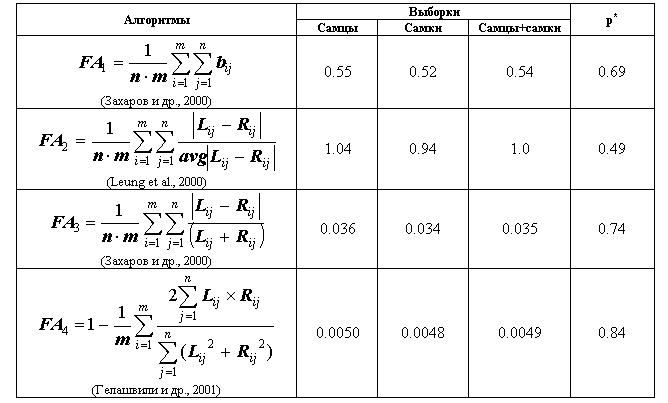
Примечание: p* – уровень значимости различий при сравнении выборок самцов и самок по критерию Манна-Уитни.
Полученные значения индексов позволяют провести сравнение выборок. Результирующие значения, характеризующие особь, заведомо отклоняются от нормального распределения, поэтому для сравнения был использован непараметрический критерий Манна-Уитни. Различий по уровню ФА между самцами и самками не обнаружено, что подтверждает ранее полученные результаты об отсутствии гендерных различий по отдельным признакам (табл. 8). Данный вывод позволяет рекомедовать объединение самцов и самок в единую выборку при проведении дальнейших исследований.
ЛИТЕРАТУРА
Аветисов В.А., Гольданский В.И. Физические аспекты нарушения зеркальной симметрии биоорганического мира // Успехи физических наук. 1996. Т. 166. № 8.
С. 873-891.
Айвазян С.А., Енюков И.С., Мешалкин Л.Д. Прикладная статистика. Основы моделирования и первичная обработка данных. М.: Финансы и статистика, 1983. 471 с.
Боровиков В.П. STATISTICA: искусство анализа данных на компьютере. Для профессионалов. СПб.: Питер, 2001а. 656 с.
Боровиков В.П. Программа STATISTICA для студентов и инженеров. М.: Компьютер-Пресс, 2001б. 301 с.
Вейль Г. Симметрия. М.: Наука, 1968. 191 с.
Гелашвили Д.Б. Еще раз о стабильности развития (по поводу статьи М. Козлова, "Заповедники и Национальные парки", № 36) // Заповедники и Национальные парки. 2002.
№ 37-38. С.45.
Гелашвили Д.Б., Краснов А.К., Логинов В.В. и др. Методологические и методические аспекты мониторинга здоровья среды государственного природного заповедника "Керженский" // Труды ГПЗ "Керженский". Н. Новгород. 2001. Т.1. С.287-325.
Гиляров М.С. О функциональном значении симметрии организмов // Зоол. журн. 1944. Т. 23. № 5. С. 213-215.
Гланц С. Медико-биологическая статистика. М.: Практика, 1998. 459 с.
Гродницкий Д.Л. Логика и неопределенность морфологических объяснений (принцип минимальных изменений в эволюции) // Журн. общей биологии. 1998. Т. 59. № 6.
С.606-620.
Жданова Н.П. Анализ фенотипической изменчивости при оптимальных и неоптимальных условиях развития в эксперименте и в природных условиях на примере прыткой ящерицы (Lacerta agilis L.): Автореф. дис. …канд. биол. наук. М.: ИБР РАН, 2003. 23 с.
Захаров В. М. Асимметрия животных. М.: Наука, 1987. 216 с.
Захаров В.М. Онтогенез и популяция (стабильность развития и популяционная изменчивость) // Экология. 2001. №3. С. 177-191.
Захаров В. М., Баранов А. С., Борисов В. И. и др. Здоровье среды: методика оценки. Оценка состояния природных популяций по стабильности развития: методологическое руководство для заповедников. М.: Центр экологической политики России, 2000. 66 с.
Кожара А. В. Структура показателя флуктуирующей асимметрии ![]() и его пригодность для популяционных исследований // Биологические науки. 1985. № 6. С. 100-103.
и его пригодность для популяционных исследований // Биологические науки. 1985. № 6. С. 100-103.
Лима-де-Фариа А. Эволюция без отбора: Автоэволюция – формы и функции. М.: Мир, 1990. 455 с.
Марченко А.О. Реализация морфогенетического потенциала растительных организмов: калибровочный подход // Журн. общей биологии. 1999. Т. 60. № 6. С. 654-666.
Реброва О.Ю. Статистический анализ медицинских данных. Применение пакета прикладных программ STATISTICA. М., МедиаСфера, 2002. 312 с.
Стабильность развития: еще раз о мнимой и реальной простоте методики / Коллектив авторов // Заповедники и Национальные парки. 2002. № 36. С.23-25.
Урманцев Ю.А. Симметрия природы и природа симметрии. М.: Мысль, 1974. 229 с.
Graham J. H., Emlen J. M., Freeman D. C., Leamy L. J., Kieser J. A. Directional asymmetry and the measurement of developmental instability // Biol. J. Lin. Soc. 1998. V. 64. P. 1-16.
Kozlov M.V., 2003. Are fast growing birch leaves more asymmetrical // Oikos. V. 101. № 3. P. 654-658.
Leung, B., Forbes M. R., Houle D. Fluctuating asymmetry as a bioindicator of stress: Comparing efficacy of analyses involving multiple traits. // American Naturalist. 2000. V. 155.
P. 101-115.
Martel J., Lempa K., 2003. A reply to Kozlov // Oikos. 2003. V. 101. № 3. P. 659-660.
Palmer A. R., Strobeck C. Fluctuating asymmetry analysis revisited // Developmental instability (DI): causes and consequences. M. Polak, ed. Oxford University Press, New York, 2003. (доступно в интернет: http://www2.biology.ualberta.ca/palmer/pubs/03BookChapt/ P&S_Chapter.pdf)
Palmer A. R., Strobeck C. Fluctuating asymmetry: measurement, analysis, patterns // Ann. Rev. Ecol. Syst. 1986. V. 17. P. 391-421.
Van Dongen S. How repeatable is the estimation of developmental stability by fluctuating asymmetry? // Proc. R. Soc. Lond. B. 1998. V. 265. P. 1423-1427.
Whitlock M. The heritability of fluctuating asymmetry and genetic control of developmental stability // Proc. R. Soc. Lond. B. 1996. V. 263. P. 849-854.
Whitlock M. The repeatability of fluctuating asymmetry: a revision and extension // Proc. R. Soc. Lond. B. 1998. V. 265. P. 1429-1431.