| Дальше
|
Назад |
Конец |
Список
|
7. Нелинейность реакции наземных экосистем на токсическую нагрузку
7.1. Дозовые зависимости для реакции наземных экосистем на токсическую нагрузку
Постановка задачи построения дозовых зависимостей для экосистемных параметров идет из классической токсикологии, где они – центральное звено в анализе действия неблагоприятных факторов на организм. С развитием экологической токсикологии была высказана идея о необходимости построения зависимостей доза–эффект для экосистемы в целом. В них “дозой” выступает величина поступления поллютантов в экосистему или их депонирования, а “эффектом” – параметры экосистемного или популяционного уровней. Именно на основе анализа таких зависимостей, в рамках развиваемой нами концепции экологического нормирования, возможно нахождение величин критических нагрузок. Однако несмотря на важность анализа дозовых зависимостей на экосистемном уровне, на настоящий момент имеется немного попыток их построения. Основная причина этого – необходимость модификации традиционной схемы работ так, чтобы были выполнены два условия: 1) достаточно большое количество пробных площадей однородно “покрывает” весь градиент нагрузки; 2) имеется информация о величине токсической нагрузки в каждой учетной точке. Выполнение этих условий при анализе многих компонентов экосистем – достаточно сложная задача.
При построении дозовых зависимостей мы использовали данные по трем районам, однако наиболее представительный материал собран только для одного (район СУМЗа) (Таблица 2). Во всех случаях были использованы интегральные параметры состояния отдельных компонентов: для древостоя – запас и густота, сомкнутость полога, доля сухостоя по запасу, плотность всходов и подроста; для травяно–кустарничкового яруса и луговой растительности – общая фитомасса, видовое богатство, видовая насыщенность, сходство с фоновым сообществом по видовой структуре, доля в фитомассе индикаторных групп (злаки, хвощи, разнотравье); для мохового яруса – проективное покрытие, видовое богатство (эпигейные, эпифитные и эпиксильные виды), сходство с фоновым сообществом по видовому списку, обилие доминирующих видов и родов; для почвы – параметры почвенного поглощающего комплекса (содержание обменного кальция и магния, степень насыщенности основаниями), сходство с фоновым профилем по оструктуренности горизонтов и выраженности техногенно–обусловленных почвообразовательных процессов; для лесной подстилки – мощность; для эпифитных лишайников – видовое богатство, видовая насыщенность, проективное покрытие на основании ствола и на высоте 1.3 м, сходство с фоновым сообществом по видовому списку, высота поднятия по стволу доминирующего вида; для почвенного микробоценоза – актуальная и потенциальная скорости деструкции целлюлозы, целлюлазная активность, интенсивность накопления аминокислот; для почвенной мезофауны – общая плотность населения, доля индикаторных групп (сапрофаги, дождевые черви, энхитреиды, элатериды), сходство таксономической и трофической структуры с фоновым сообществом.
Таблица 2. Объем материала, использованного для построения и анализа зависимостей доза – эффект для реакции наземных экосистем
|
Характеристика работ |
Район действия медеплавильного завода |
||
|
Среднеуральского |
Красноуральского |
Кировградского |
|
|
Годы сбора материала |
1989 – 2001 |
1989, 2000 |
1995 |
|
Компоненты экосистемы |
древесный ярус (6)*, травяно–кустарничковый ярус (5), луговая растительность (5), моховой ярус (6), эпифитные лишайники (2 – 3), лесная подстилка (1), почва (4), почвенная мезофауна (6), почвенный микробоценоз (4) |
травяно–кустарничковый ярус (4), эпифитные лишайники (4), почвенная мезофауна (3) |
древесный ярус (3), травяно–кустарничковый ярус (5), моховой ярус (3), эпифитные лишайники (6), лесная подстилка (1), почвенный микробоценоз (3) |
|
Количество: |
|||
|
пробных площадей |
от 14 до 208 |
16 – 20 |
15 |
|
информационных массивов |
42 |
3 |
5 |
|
зависимостей доза – эффект |
119 |
11 |
20 |
Примечание. * – в скобках приведено количество интегральных параметров для компонента.
Во всех случаях при аппроксимации зависимостей регрессионными уравнениями в качестве учетной единицы использована пробная площадь, т.е. средние значения как для параметра биоты, так и для величины токсической нагрузки. Для получения оценок нагрузки использованы 3 – 5 физически усредненных образцов депонирующих сред (каждый из которых составлен из 5 индивидуальных), а для параметров биоты – общепринятое для конкретных объектов количество индивидуальных измерений (15 проб 50х50 см для травяно – кустарничкового яруса, 8 проб 50х50 см для луговой растительности, 10 деревьев–форофитов для лихеносинузий, 15 – 30 прикопок для лесной подстилки, 10 проб 1х1 м для мохового яруса, сплошной перечет деревьев на пробной площади 25х25 м, 5 проб 1х1 м для подроста, 10 – 40 проб 20х20 см для почвенной мезофауны, 10 – 20 проб для биологической активности почвы).
Для района СУМЗа, помимо более широкого охвата компонентов экосистем по сравнению с районами КМК и КМЗ, мы имели возможность разделять материал на подвыборки (информационные массивы) в зависимости от определенных критериев (положение в рельефе, последовательные годы наблюдений и др.), что позволило сравнить дозовые зависимости для разных вариантов биотопов и оценить их межгодовую изменчивость. Количество точек в информационных массивах, использованных для построения и анализа дозовых зависимостей было не менее 14 и, как правило, составляло 25 – 30. В общей сложности проанализировано 150 дозовых зависимостей для интегральных параметров состояния наземных экосистем по 50 информационным массивам.
7.1.1. Сравнительный анализ дозовых зависимостей для реакции разных компонентов экосистем
В большинстве случаев дозовые зависимости для реакции компонентов наземных экосистем имели “классическую” форму S–образной кривой (Рисунок 10). Эта было справедливо для всех рассмотренных районов и большинства показателей и позволило удовлетворительно аппроксимировать зависимости логистическими уравнениями регрессии (при этом доля объясняемой дисперсии в большинстве случаев лежала в пределах 70 – 90%). В тех случаях, когда наблюдались отклонения от S–образной формы, они чаще всего касались редукции верхнего (или нижнего для восходящих кривых) плато, что было связано с недостаточным количеством точек в области низких нагрузок. Другой вариант – дозовая зависимость представляла собой комбинацию двух логистических кривых (см. далее Рисунок 15).
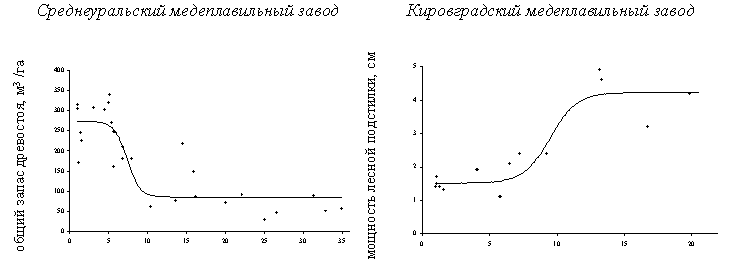
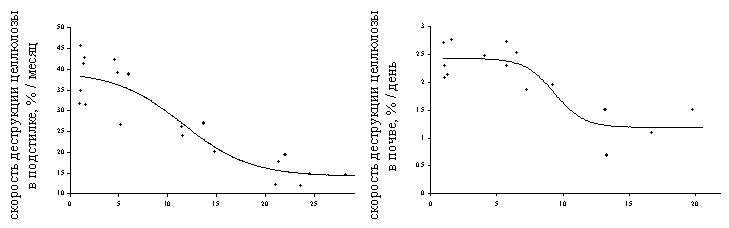
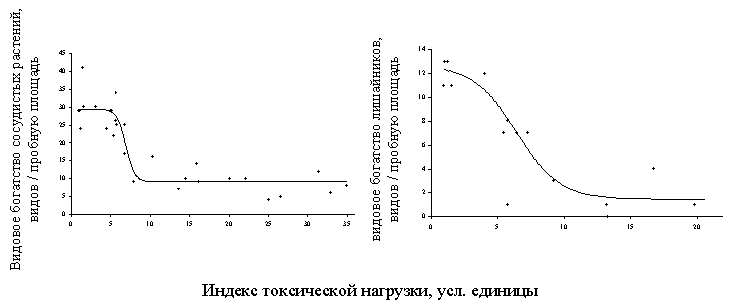
Рисунок 10. Примеры зависимостей доза – эффект для параметров состояния наземных экосистем в районе Среднеуральского и Кировградского медеплавильных заводов.
Один из важнейших результатов анализа дозовых зависимостей заключается в их ярко выраженной ступенчатости: для большинства экосистемных параметров реакция на нагрузку нелинейна – существует два относительно стабильных уровня, соответствующих фоновому и импактному состояниям, с очень резким переходом между ними. Параметр, характеризующий резкость ступени – доля градиента (от его общей величины), которая приходится на область перехода. Для большинства рассмотренных показателей этот параметр не превышает 5 – 10 % (Рисунок 11), что позволяет охарактеризовать переходы как очень “быстрые”, происходящие по типу триггера. Лишь небольшая часть параметров изменяется плавно (доля участка градиента между верхней и нижней критическими точками составляет 25 – 40 %). Другими словами, экосистема реагирует на увеличение загрязнения не постепенными изменениями, а по закону “все или ничего”. Существование порога в реакции экосистемы, т.е. области нагрузок, при которых не обнаруживается существенных изменений, есть проявление феномена устойчивости, наличия эффективных механизмов регуляции. Соответственно подпороговые значения нагрузок – это оценка “запаса гомеостатичности” экосистемы, в пределах которого воздействия на нее допустимы. Именно нелинейность реакции придает объективный статус определяемым критическим нагрузкам и вводимым экологическим нормативам, соответствующим началу выхода из стабильного состояния. Если бы зависимость доза–эффект имела вид прямой линии или плавной кривой, такого объективного критерия не существовало, что потребовало бы поиска других подходов к нормированию, либо вынудило устанавливать нормативы произвольно. Таким образом, именно пороговость в дозовой зависимости существенно упрощает решение задачи экологического нормирования.
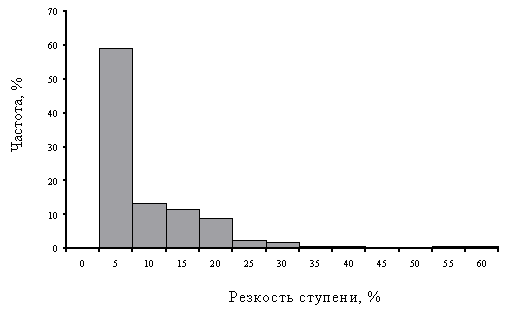
Рисунок 11. Частотное распределение резкости перехода от фонового к импактному состоянию для всех рассмотренных интегральных показателей наземных экосистем (по оси абсцисс – доля от максимального значения нагрузки, верхняя граница интервала).
Сопоставление критических нагрузок (соответствуют абсциссам верхних критических точек для ниспадающих кривых и нижних – для восходящих) для рассмотренных компонентов экосистем показывает их значительное различие в изученных районах (Рисунок 12) (для целей сравнения критические нагрузки выражены как доля от максимальной нагрузки). В районе КМЗ экосистемы начинают изменяться при относительно более высоких уровнях нагрузки, чем в районах СУМЗа и КМК. Скорее всего, это связано с малой подвижностью тяжелых металлов в почве в данном районе из-за подщелачивания кальцийсодержащей пылью.
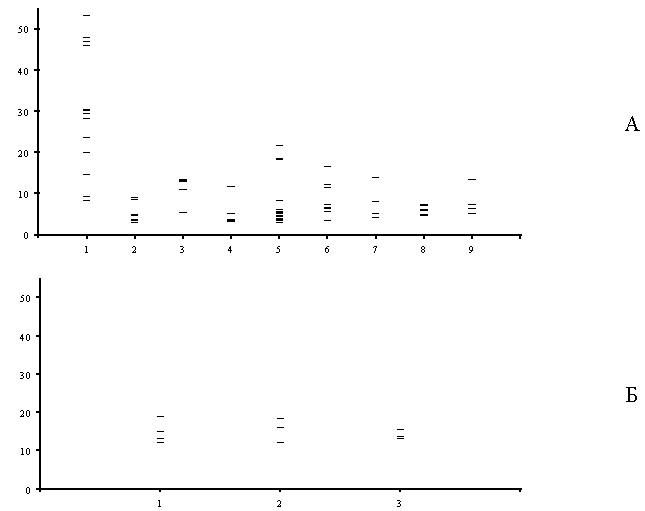
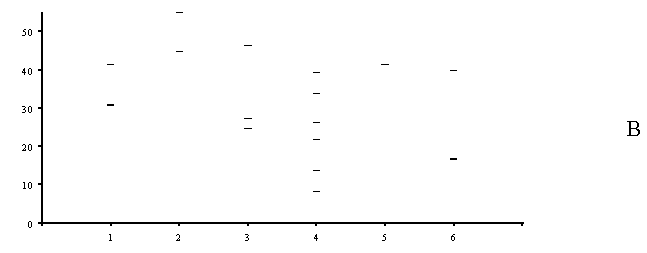
Рисунок 12. Относительное значение критических нагрузок для разных показателей состояния наземных экосистем (по оси ординат – доля от максимальной нагрузки, %):
А – район СУМЗа: 1 – древесный ярус, 2 – травяно – кустарничковый ярус лесных сообществ, 3 – травостой луговых сообществ, 4 – моховой ярус, 5 – эпифитные лишайники, 6 – лесная подстилка, 7 – почва, 8 – почвенная фауна, 9 – почвенный микробоценоз.
Б – район КМК: 1 – травяно – кустарничковый ярус, 2 – эпифитные лишайники, 3 – почвенная фауна.
В – район КМЗ: 1 – древесный ярус, 2 – травяно – кустарничковый ярус, 3 – моховой ярус, 4 – эпифитные лишайники, 5 – лесная подстилка, 6 – почвенный микробоценоз.
Для района СУМЗа часть компонентов (древесный ярус, лесная подстилка) демонстрирует широкий спектр критических нагрузок (от 5 до 50%). В эту же группу попадают и эпифитные лишайники, хотя их обычно относят к наиболее чувствительным индикаторам атмосферного загрязнения. Для других компонентов критические нагрузки лежат очень “кучно”, однако пока не ясно в какой степени это связано с полнотой описания этих компонентов. Минимальные критические нагрузки для большинства компонентов имеют близкие значения (в диапазоне от 3 до 10%). Данный результат можно интерпретировать как то, что нагрузки, при которых начинают изменяться наиболее чувствительные показатели примерно одинаковы для всех компонентов экосистем. Исключение составляет луговая растительность, изменение которой регистрируется при относительно более высоких нагрузках.
7.1.2. Межгодовая изменчивость дозовых зависимостей
Вопрос о стабильности параметров дозовых зависимостей во времени важен в плане надежности определения величин критических нагрузок: если межгодовая изменчивость велика, параметры дозовых зависимостей, определенные в конкретный год, рискованно экстраполировать на другие периоды. Для анализа межгодовой изменчивости было выбрано два объекта в районе СУМЗа – травяно–кустарничковый ярус лесных сообществ (елово–пихтовые леса) и травостой луговых сообществ.
Для изучения реакции травяно–кустарничкового яруса было заложено 28 постоянных пробных площадей, на которых по единой методике в течение 5 лет (с 1998 по 2002 годы) проводились описания растительности. Дозовые зависимости были построены как для функциональных параметров (общая фитомасса), так и для структурных (параметры разнообразия, доля в фитомассе индикаторных групп, сходство видовой структуры с фоновым сообществом). Дозовые зависимости для общей фитомассы травяно–кустарничкового яруса лесных сообществ существенно различаются по годам (Рисунок 13, Таблица 3). Анализируя данные 2000 г. можно прийти к заключению о почти полном отсутствии зависимости общей фитомассы от величины нагрузки. Именно к такому выводу мы пришли ранее (Воробейчик, Хантемирова, 1994), рассматривая такую же зависимость по данным, полученным в 1989 г. на тех же самых пробных площадях. С другой стороны, зависимости по данным 1999 г. и 2002 годов имеют “классическую” S–образную форму.
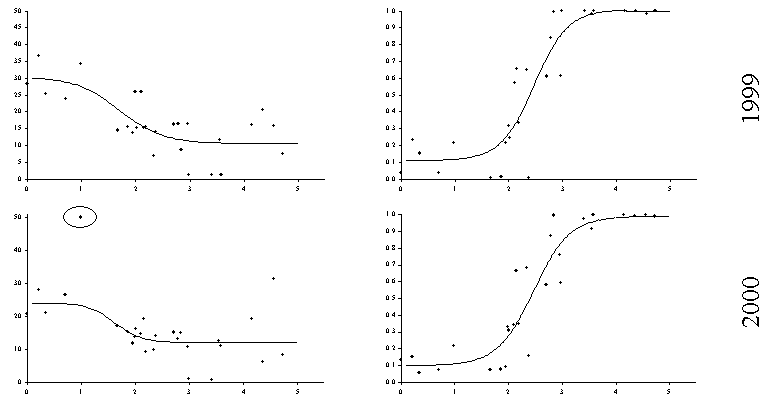
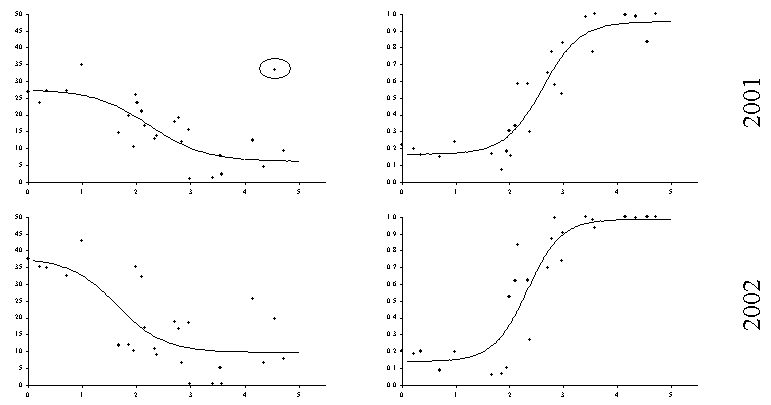
Рисунок 13. Межгодовая изменчивость дозовых зависимостей для реакции травяно–кустарничкового яруса лесных сообществ. Токсическая нагрузка оценена по содержанию обменных форм тяжелых металлов в лесной подстилке. Эллипсом обведены точки, которые были исключены при аппроксимации зависимостей уравнениями регрессии.
Таблица 3. Параметры дозовых зависимостей для реакции травяно–кустарничкового яруса лесных сообществ за 5 последовательных лет
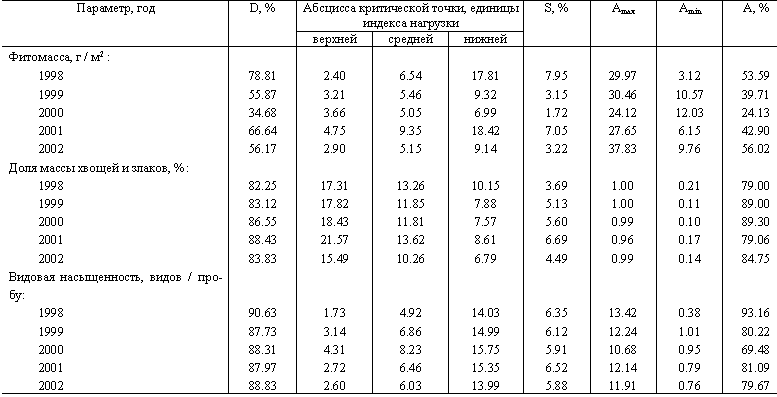
Примечание. D – доля объясняемой логистическим уравнением дисперсии, S – участок градиента с быстрыми изменениями параметра (доля от общей величины градиента), Amax и Amin – верхняя и нижняя асимптоты логистической функции, A – относительная амплитуда изменений (разница между верхней и нижней асимптотами, отнесенная к максимальному значению параметра, зарегистрированному за все 5 лет наблюдений).
Вариант отсутствия зависимости общей фитомассы от величины токсической нагрузки легко объяснить наличием компенсаторных реакций: подавление чувствительных видов компенсируется увеличением обилия более толерантных к действию загрязнения. В нашем случае такими толерантными группами видов, увеличивающими обилие по мере приближения к источнику выбросов, оказываются хвощи (хвощ лесной и луговой) и злаки (щучка дернистая и полевица). Сравнение данных за несколько лет свидетельствует, что компенсаторные реакции включаются не всегда, что скорее всего связано с межгодовыми различиями гидро–термических условий.
В отличие от общей фитомассы, дозовые зависимости для структурных параметров травяно–кустарничкового яруса лесных сообществ демонстрируют более высокую стабильность. Учитывая характерную для экологических данных вариабельность, связанную как с природной гетерогенностью популяций и сообществ, так и с неизбежными ошибками измерений, данный результат представляется важным в методическом отношении.
Аналогичные результаты были получены для луговых сообществ, анализ которых был выполнен на 20 постоянных пробных площадей, на которых описания проводились в течение 4 лет (с 1999 по 2002 годы).
7.1.3. Экотопические различия дозовых зависимостей
Различия параметров дозовых зависимостей между разными вариантами экотопов были рассмотрены на примере двух объектов – лесной подстилки и синузий эпифитных лишайников (видовое разнообразие на березе), информация о которых собрана на 208 пробных площадях в районе СУМЗа (1995, 1996 и 1998 гг.). Такое количество площадок позволило получить представительные выборки, характеризующие разные варианты экотопов в зависимости от положения в рельефе, состава древостоя, типа почвы, освещенности и увлажненности (две последние характеристики получены на основе геоботанических описаний травяно – кустарничкового яруса с использованием экологических шкал Элленберга).
Характер экотопа существенно влияет на величину критической нагрузки для мощности лесной подстилки, которую мы рассматриваем как интегральный параметр интенсивности деструкционных процессов в лесной экосистеме, детерминируемых активностью почвенной биоты (Рисунок 14). При сравнении биотопов, различающихся положением в рельефе, прослеживается четкая закономерность: величина критической нагрузки увеличивается в ряду аккумулятивные – транзитные – элювиальные ландшафты. Разница между критическими нагрузками для транзитных ландшафтов по сравнению с аккумулятивными и элювиальных по сравнению с транзитными составляет почти 2 раза. Дополнительную информацию о значительности обнаруженных различий дает сопоставление абсцисс других критических точек. Мощность подстилки в транзитных ландшафтах начинает изменяться при таких уровнях нагрузки, которые в аккумулятивных ландшафтах уже наполовину изменили фоновый уровень. Когда же в аккумулятивных ландшафтах изменения завершены и мощность подстилки стабилизировалась на верхнем плато, в элювиальных ландшафтах регистрируется только ее начальное увеличение. Интересно то, что экотопические различия не влияют на форму дозовых кривых, а только меняют их количественные параметры (абсциссы критических точек). В методическом плане важно также, что контрастные варианты биотопов очень мало или вовсе не различаются по начальным (фоновым) и конечным (импактным) значениями мощности подстилки.
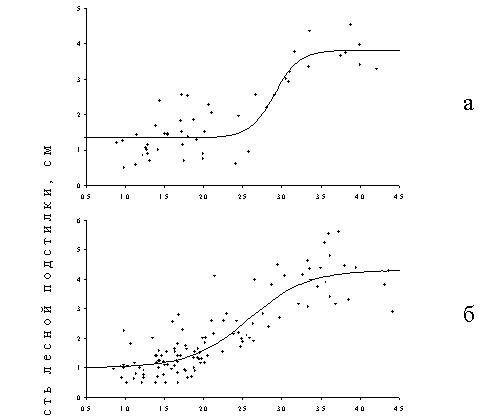
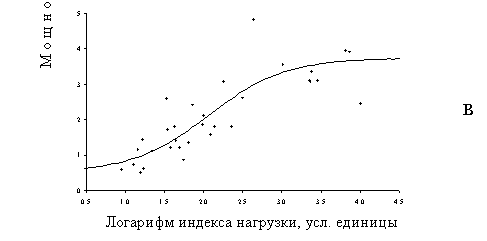
Рисунок 14. Зависимости доза–эффект для изменения мощности лесной подстилки в разных элементах рельефа: а – элювиальных, б – транзитных, в – аккумулятивных. Токсическая нагрузка оценена по содержанию подвижных форм тяжелых металлов в лесной подстилке.
Среди нескольких гипотез, которые могут объяснить феномен меньшей устойчивости почвенной биоты в аккумулятивных ландшафтах, наиболее правдоподобной нам представляется гипотеза синергизма действия поллютантов и неблагоприятного для почвенной биоты сочетания естественных факторов (избыточное увлажнение, формирующее в почве анаэробные условия), что чаще имеет место в аккумулятивных ландшафтах по сравнению с элювиальными и транзитными.
В большинстве случаев дозовые зависимости для реакции эпифитных лишайников хорошо аппроксимировала комбинация двух логистических кривых (Рисунок 15). Соответственно, выделяются три относительно стабильных состояния – фоновое, промежуточное и импактное, а также два переходных между стабильными. Наиболее существенные изменения параметров лихеносинузий происходят при переходе от промежуточного состояния к импактному, тогда как переход от фонового состояния к промежуточному выражен значительно менее контрастно (существенно меньшая разница между асимптотами для верхней кривой по сравнению с нижней). В экотопах со слабым освещением эпифитные лишайниковые синузии выдерживают более сильную токсическую нагрузку по сравнению с сильно освещенными. Кривые для средне освещенных экотопов занимают промежуточное положение (за исключением верхней кривой для видового богатства, которая демонстрирует высокую чувствительность в низкодозовой части градиента; возможная причина этого – малое количество точек в этой части градиента, не позволяющее провести надежное определение критической нагрузки). Чувствительность лишайниковых синузий также различается в разных элементах рельефа: сообщества элювиальных ландшафтов выдерживают более сильную нагрузку по сравнению с транзитными и аккумулятивными. Сравнение местообитаний с разной степенью увлажнения демонстрирует максимальную чувствительность к токсической нагрузке в сильно увлаженных экотопах.
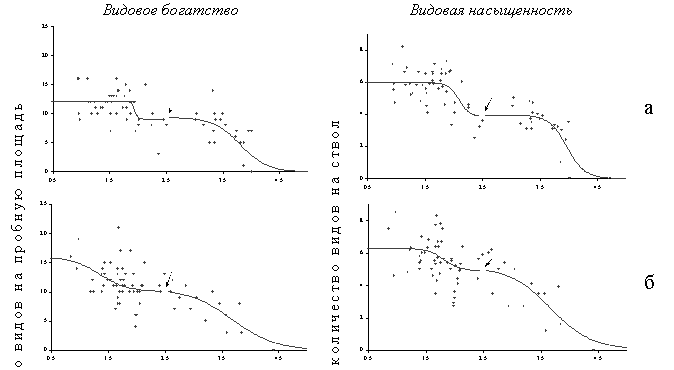
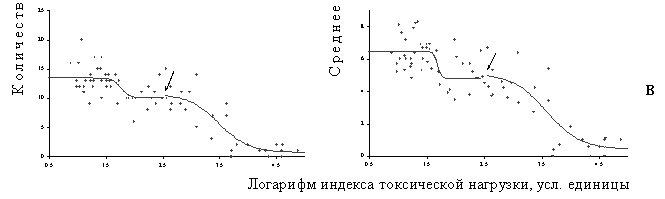
Рисунок 15. Зависимости доза – эффект для параметров эпифитных лишайниковых синузий в разных вариантах биотопов: а – минимальная освещенность, б – средняя освещенность, в – максимальная освещенность. Токсическая нагрузка оценена по содержанию подвижных форм тяжелых металлов в лесной подстилке. Стрелкой обозначена граница между графиками двух логистических функций.
Скорее всего, обнаруженные экотопические различия в реакции эпифитных лишайников связаны с тем, что разные виды в одних вариантах биотопов выдерживают более высокие нагрузки, а в других – более низкие (что связано с физиологическими, анатомическими или морфологическими особенностями). Дозовые же зависимости для таких интегральных параметров, как видовое богатство и видовая насыщенность отслеживают баланс между количеством толерантных и чувствительных видов для конкретного варианта экотопа.
Важно отметить существенное сходство результатов по экотопическим различиям дозовых зависимостей для столь разных объектов, как лесная подстилка и эпифитные лишайники. М.Р. Трубина (2002), выполнившая на этом же материале предварительный анализ дозовых зависимостей для реакции травяно–кустарничкового яруса, также пришла к выводу о значительных экотопических различиях. Все это можно рассматривать как то, что данная закономерность имеет значительный уровень общности и, скорее всего, будет справедлива для большинства компонентов наземных экосистем. В методическом плане полученные результаты свидетельствуют о важности дифференциации экологических нормативов в зависимости от экотопических условий.
7.2. Роль пространственной неоднородности среды в механизмах реакции экосистем на токсическую нагрузку
Вопрос о механизмах возникновения нелинейности в реакции биоты на токсическую нагрузку распадается на три составляющие, соответствующие трем участкам “классической” дозовой кривой, а именно: “почему наблюдается эффект порога?”, “почему наблюдается эффект триггера?”, “почему в области больших нагрузок уже не происходит существенных изменений?”. Основной из них – вопрос о триггерном эффекте. Фактически, существование нелинейности означает, что в небольшой относительно общей величины градиента загрязнения переходной зоне (т.е. в очень узком интервале токсических нагрузок) резко увеличено разнообразие наблюдаемых состояний экосистем. Другими словами при одном и том же уровне нагрузки могут быть зарегистрированы как почти фоновые, так и почти импактные варианты состояния. Такое переформулирование проблемы из терминов “реакции” в термины “разнообразия состояний” позволяет выдвинуть несколько рабочих гипотез для объяснения триггерного эффекта:
Сформулированные гипотезы могут быть экспериментально проверены, что частично было выполнено нами для района СУМЗа.
7.2.1. Пространственное варьирование микроклиматических параметров
Было рассмотрено пространственное варьирование освещенности под пологом леса, влажности почвы и подстилки, температуры почвы на глубине 5 и 10 см (Рисунок 16). Работы выполнены в трех зонах токсической нагрузки (фоновая, буферная, импактная) в двух пространственных масштабах – единиц метров (линии из 80 или 100 точек опробования, расположенных через 30 или 50 см) и сотен метров (25 пробных площадок 10х10 м, относительно равномерно размещенных на участке размером порядка 500х500 м). Всего выполнено 3150 измерений освещенности, 3960 – температуры почвы, 1100 – влажности почвы и подстилки.
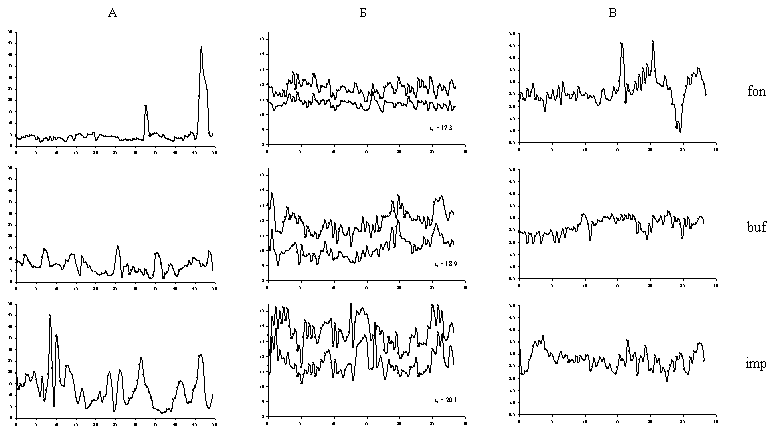
Рисунок 16. Пространственное варьирование микроклиматических параметров в регулярных линиях в фоновой (fon), буферной (buf) и импактной (imp) зонах нагрузки: А – освещенность под пологом леса, % от открытого места, Б – температура почвы (оС) на глубине 5 см (светлые точки) и 10 см (темные точки), ta – средняя температура воздуха за период измерений, В – влажность подстилки (г воды / г сухой массы). По оси абсцисс – расстояние в м.
Увеличение варьирования микроклиматических параметров исходно связано с разрушением древостоя и индуцированного этим осветления местообитаний: средняя для зоны освещенность на фоновой территории составляет 11.5% от открытого места, в буферной зоне 13.7%, в импактной – 15.6%. Пространственная вариабельность освещенности увеличивается под действием токсической нагрузки: в масштабе сотен метров коэффициент вариации средней на площадку освещенности возрастает в два раза – от 22.8% на фоновой территории до 41.3% в буферной и 38.9% в импактной зонах. В масштабе единиц метров на загрязненной территории сглаживаются различия между пологом леса и окнами, которые очень контрастны на фоновой территории. Если из линий на фоновой территории “удалить” хорошо диагностируемые окна древостоя, разница между зонами по пространственной вариабельности в масштабе единиц метров становится еще более контрастной, чем в масштабе сотен метров: коэффициент вариации возрастает от 22.8 – 38.5 % на фоновой территории до 44.1 – 45.1 % на буферной и 54.2 –76.0 % на импактной.
Разница между зонами по пространственному варьированию температуры регистрируется только для теплых погодных условий: в масштабе единиц метров интерлимитный размах температуры на глубине 5 см в фоновой зоне составляет 2.1 оС, в буферной – 3.5 оС, импактной – 4.2 оС (среднеквадратическое отклонение – 0.4, 0.8 и 0.9 оС соответственно). Для холодных погодных условий различия между зонами по пространственному варьированию температуры практически отсутствуют.
Пространственное варьирование влажности почвы и подстилки не демонстрирует четких тенденций изменения в градиенте загрязнения. В одном случае (масштаб единиц метров) было зарегистрировано уменьшение варьирования влажности подстилки (коэффициент вариации снизился с 21.7 % на фоновой территории до 12.6 – 13.9 % в буферной и импактной зонах). В другом – наблюдалась противоположная ситуация: в масштабе сотен метров коэффициент вариации влажности подстилки увеличился с 28.5% на фоне до 33.3 – 36.3% в буферной и импактной зонах (для почвы разница была еще выше – 20.1% на фоновой территории и 40.7% на импактной).
В целом, пространственное варьирование микроклиматических параметров существенно уступает варьированию содержания металлов и кислотности в условиях токсических нагрузок (см. раздел 7.2.2). Следовательно, данный фактор нельзя считать основным в объяснении механизмов триггерного эффекта.
7.2.2. Пространственная неоднородность химического загрязнения природных депонирующих сред
Изменение в градиенте нагрузки пространственного варьирования загрязненности было исследовано для нескольких депонирующих сред (снег, лесная подстилка, почва) и в широком диапазоне масштабов – от десятков сантиметров – единиц метров до сотен метров – единиц километров (Таблица 4). Помимо традиционных мер варьирования мы использовали относительный размах – отношение интерлимитного размаха в конкретной зоне к интерлимитному размаху по всему градиенту нагрузки (характеризует долю от всего диапазона возможного варьирования).
Таблица 4. Объем материала, использованного для анализа пространственной неоднородности химического загрязнения депонирующих сред
|
Характеристика материала |
Депонирующая среда |
||
|
снег |
лесная подстилка |
почва |
|
|
Схема отбора |
сетка |
сетка, линия, “равномерно – случайное” |
сетка, линия, “равномерно – случайное” |
|
Пространственный масштаб |
10х10 м (2 м)* 100х100 м (20 м) 5х20 км (1 км) |
1х1 м (0.2 м) |
1х1 м (0.2 м) |
|
Регистрируемые параметры |
Масса пылевых частиц, рH, водо– и кислоторастворимые формы Cu, Cd, Pb, Zn |
pHводный; подвижные и обменные формы Cu, Cd, Pb, Zn, Fe |
pHводный; подвижные и обменные формы Cu, Cd, Pb, Zn, Fe |
|
Количество : |
|||
образцов |
600 |
2 350 |
1 900 |
измерений |
6 600 |
25 500 |
13 640 |
Примечание. * – в скобках приведено расстояние между точками отбора.
Пространственное варьирование кислотности относительно стабильно в градиенте загрязнения. Для масштаба десятков сантиметров – единиц метров размах кислотности подстилки во всех зонах нагрузки равен 0.5 – 1.5 единицы рН, что составляет 12 – 45% от всего диапазона возможного варьирования (для почвы 0.2 – 1.0 единицы рН и 6 – 27% соответственно). Для масштаба сотен метров – единиц километров варьирование существенно выше: и для подстилки, и для почвы размах кислотности лежит в пределах 1.1 – 3.3 единицы рН, что составляет 30 – 75% от всего диапазона возможного варьирования. Коэффициент вариации концентраций иона водорода во всех зонах нагрузки очень велик: для масштаба единиц километров он лежит в пределах 60 – 210%.
Высокое пространственное варьирование кислотности важно учитывать при анализе реакции биоты на токсическую нагрузку, поскольку именно от данного параметра зависит протекание многих процессов, модифицирующих токсичность почвы и подстилки. При возрастании концентрации ионов водорода в почвенном растворе снижается степень насыщенности основаниями, увеличивается содержание подвижного алюминия и гидролитическая кислотность (Рисунок 17). Наиболее резкий рост содержания подвижного Al, обуславливающего эндогенную токсичность почвы и подстилки для растений, наблюдается при рН > 5. Полученная зависимость не выходит за рамки представлений о влиянии кислотности на растворимость соединений алюминия: известно, что в широком диапазоне от слабокислых до слабощелочных сред алюминий представлен труднорастворимым гидроксидом Al(OH)3, а граница перехода алюминия в раствор в кислых почвах как раз и пролегает на уровне 5 единиц рН. Важно отметить, что характер зависимости и диапазон изменений параметров очень сходен во всех зонах нагрузки. Это означает, что очень высокие и очень низкие концентрации подвижного алюминия, в зависимости от кислотности, могут наблюдаться во всех зонах нагрузки. Сказанное справедливо и для такого параметра почвенного поглощающего комплекса, как степень насыщенности основаниями, в значительной степени детерминирующего буферность почв по отношению к тяжелым металлам.
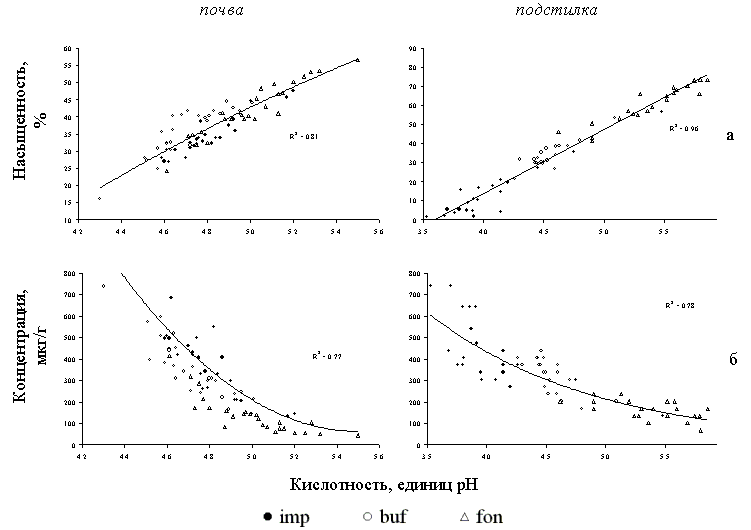
Рисунок 17. Зависимость степени насыщенности основаниями (а), содержания подвижного алюминия (б) и гидролитической кислотности (в) от актуальной кислотности (рН водный) для образцов почвы и подстилки из фоновой (fon), буферной (buf) и импактной (imp) зон токсической нагрузки. Субстраты отобраны по сетке 100х100 м (по 25 образцов в каждой зоне); учетная единица – образец.
Обменные формы металлов (экстрагирование 0.05 М раствором хлорида кальция) дают представление о той их части, которая доступна для биоты и, следовательно, может оказывать токсическое действие. Отношение “обменные формы / подвижные формы” (что близко к доле обменных форм от валового содержания) также существенно зависит от актуальной кислотности (Рисунок 18). Зависимость близка к экспоненциальной: наиболее сильное увеличение доли обменных форм происходит при снижении кислотности в диапазоне 5.5 – 5.0 единиц рН. Существуют различия в характере связи для рассматриваемых металлов на разных участках градиента загрязнения: для кадмия и цинка эта связь наблюдается во всех зонах нагрузки, тогда как для меди и свинца – только в импактной и буферной зонах. Также наблюдаются различия в абсолютных значениях доли обменных форм: для кадмия и цинка при максимальном уровне загрязнения в зависимости от кислотности они составляют от 10 до почти 100 %, тогда как для меди – от 1 до 30 %, а свинца – до 8 %.
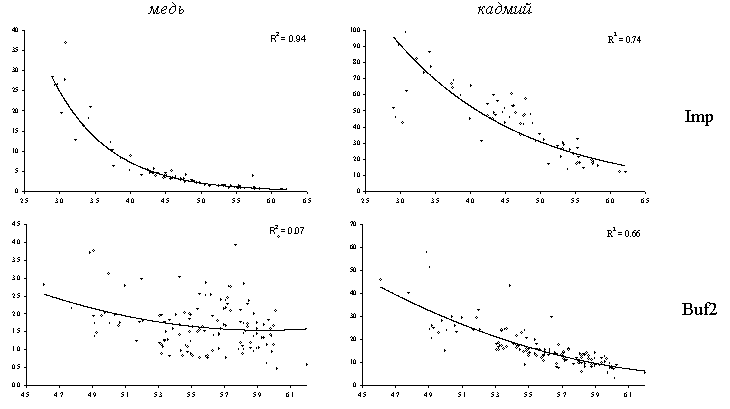
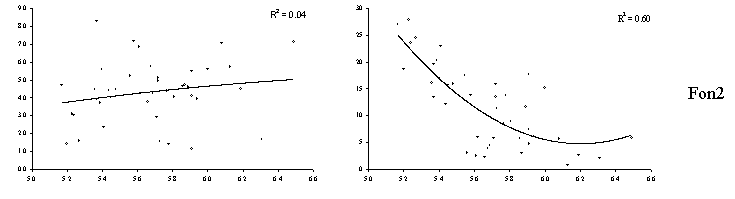
Рисунок 18. Зависимость отношения “обменные формы / подвижные формы” (%, ось ординат) от кислотности (единиц рН, ось абсцисс) лесной подстилки в разных зонах нагрузки (район СУМЗа, масштаб – 50 х 50 км) для меди и кадмия. Учетная единица – образец.
В контексте обсуждаемой проблемы модификации токсичности важно подчеркнуть, что даже при одинаковом валовом содержании в почвенный раствор могут выходить в зависимости от кислотности как очень малые, так и очень большие количества металлов. Учитывая чрезвычайно большое варьирование кислотности во всех зонах нагрузки (а в импактной и буферной зонах частотное распределение кислотности как раз покрывает “критический” для обменных форм диапазон значений рН), можно предположить, что именно это приводит к увеличению варьирования токсичности и, соответственно, росту разнообразия состояний биоты в переходной части градиента нагрузки.
Анализ пространственного варьирования содержания металлов в диапазоне масштабов от десятков сантиметров до сотен метров во всех зонах нагрузки показывает увеличение варьирования при переходе от подвижных форм к обменным и, до определенной степени, по мере увеличения площади опробования (коэффициент вариации возрастает от 10–20% до 50–80%). Пространственное варьирование и подвижных, и обменных форм выше в почве по сравнению с подстилкой (что связано, вероятно, с более разнообразными механизмами, модифицирующими содержание металлов). В большинстве случаев наблюдается следующая закономерность: пространственная вариабельность минимальна в фоновой зоне и максимальна в буферной, импактная зона занимает промежуточное положение. Даже в пределах относительно небольшого участка (линия длиной 30 м) в импактной зоне размах концентраций очень велик (составляет 63 – 81% от всего диапазона варьирования для подвижных форм и 75 – 98 % – обменных).
В масштабе единиц километров пространственное варьирование содержания металлов в почве и подстилке значительно увеличивается при переходе к импактной зоне (Таблица 5): для обменных форм коэффициент вариации по ряду элементов достигает 130–210%, а относительный размах составляет почти 100% (для подвижных форм максимальные значения коэффициента вариации также очень высокие – 90 – 100%). Существуют различия между элементами в трендах изменения пространственного варьирования обменных форм в градиенте загрязнения: коэффициент вариации для кадмия и цинка, в отличие от меди, свинца и железа, не увеличивается от фоновой зоны к импактной, а остается стабильным или даже уменьшается. Вероятно, это связано с рассмотренными выше различиями в характере связи между концентрацией рассматриваемых металлов и кислотностью в разных зонах нагрузки.
Таблица 5. Параметры пространственного варьирования концентраций обменных и подвижных форм тяжелых металлов в лесной подстилке и верхнем слое почвы в разных зонах токсической нагрузки в масштабе участка 50х50 км (всего по 620 образцов почвы и подстилки)
|
Элемент |
Коэффициент вариации, % |
Относительный размах, % |
||||||||
|
fon2 |
fon1 |
buf2 |
buf1 |
imp |
fon2 |
fon1 |
buf2 |
buf1 |
imp |
|
|
Подвижные формы |
||||||||||
|
Подстилка: |
||||||||||
Cu |
27.34 |
45.36 |
46.70 |
36.76 |
46.65 |
0.70 |
3.16 |
11.01 |
21.06 |
84.67 |
Cd |
25.41 |
30.26 |
30.23 |
24.42 |
63.02 |
4.11 |
13.22 |
18.40 |
26.83 |
94.75 |
Pb |
24.70 |
35.49 |
33.82 |
32.89 |
43.21 |
2.66 |
8.27 |
18.17 |
39.55 |
87.70 |
Zn |
24.85 |
26.87 |
27.94 |
30.74 |
54.99 |
12.08 |
21.48 |
28.66 |
37.95 |
98.82 |
|
Почва: |
||||||||||
Cu |
44.73 |
50.62 |
59.16 |
46.26 |
90.21 |
2.27 |
7.05 |
16.06 |
22.04 |
97.67 |
Cd |
65.33 |
49.23 |
61.11 |
56.40 |
65.28 |
22.99 |
33.84 |
59.16 |
95.48 |
65.17 |
Pb |
48.51 |
45.18 |
61.42 |
57.79 |
105.98 |
12.23 |
26.56 |
49.66 |
43.88 |
99.81 |
Zn |
61.36 |
56.74 |
67.18 |
58.83 |
59.42 |
30.03 |
61.06 |
86.04 |
95.55 |
73.17 |
|
Обменные формы |
||||||||||
|
Подстилка: |
||||||||||
Cu |
40.04 |
44.94 |
49.52 |
72.10 |
112.70 |
0.36 |
1.51 |
1.87 |
7.59 |
98.60 |
Cd |
66.49 |
56.88 |
54.46 |
45.72 |
56.92 |
3.33 |
10.74 |
17.87 |
31.00 |
91.12 |
Pb |
61.33 |
59.96 |
62.12 |
100.37 |
133.82 |
2.29 |
3.32 |
4.54 |
10.81 |
99.87 |
Zn |
73.79 |
50.03 |
48.64 |
35.51 |
48.21 |
6.64 |
12.46 |
24.98 |
29.22 |
84.05 |
Fe |
62.35 |
58.52 |
58.23 |
81.43 |
207.01 |
0.63 |
0.78 |
0.92 |
3.49 |
99.78 |
|
Почва: |
||||||||||
Cu |
29.24 |
48.55 |
57.03 |
69.58 |
81.33 |
0.86 |
3.19 |
5.62 |
15.28 |
98.49 |
Cd |
55.34 |
55.76 |
57.78 |
45.41 |
58.44 |
11.15 |
34.60 |
48.61 |
65.69 |
97.12 |
Pb |
53.63 |
55.96 |
58.04 |
53.05 |
128.07 |
9.25 |
22.82 |
14.65 |
14.54 |
98.89 |
Zn |
66.60 |
65.98 |
67.49 |
46.98 |
46.45 |
17.86 |
53.24 |
76.70 |
78.77 |
83.69 |
Fe |
66.70 |
69.90 |
58.95 |
92.96 |
142.35 |
3.60 |
7.81 |
6.25 |
20.02 |
99.55 |
Примечание. Учетная единица – образец. Зоны нагрузки: imp – импактная (включая техногенную пустыню, 54.8 км2), buf1 – переход между импактной и буферной (247.7 км2), buf2 – буферная (358.8 км2), fon1 – переход между буферной и фоновой (1145.8 км2), fon2 – фоновая (44.2 км2).
Таким образом, на тех участках градиента загрязнения, где наблюдается переход биоты из фонового состояния в импактное, имеет место очень высокое пространственное варьирование содержания тяжелых металлов, а также очень высокое варьирование модифицирующих токсичность параметров почвы и подстилки.
7.2.3. Модельный эксперимент: биотестирование токсичности почвы и подстилки
Прямое измерение токсичности можно выполнить с помощью биотестирования субстратов, позволяющего оценить суммарное влияние многих факторов. С целью анализа связи почвенных параметров с токсичностью мы сопоставили микромасштабное пространственное варьирование содержания разных форм металлов, кислотности и фитотоксичности, оцененной по ростовой реакции растений.
Биотестирование выполнено для лесной подстилки и почвы (по 75 образцов), отобранных в ельниках – пихтарниках, расположенных в районе СУМЗа в фоновой (20 км в западу от завода), буферной (4.5 км) и импактной (1 км) зонах токсической нагрузки. В каждой зоне образцы отбирали с площадки 100х100 м. В качестве тест–объекта для оценки токсичности использовали проростки одуванчика лекарственного. Для данного вида известно, что семенное потомство одного растения представляет собой чистую линию. Использование такого генетически однородного материала позволяет максимально сократить вариабельность, вносимую индивидуальной генетической изменчивостью. Токсичность субстрата оценивали по корневому тесту. Семена проращивали методом рулонной культуры в сосудах, содержащих суспензию подстилки или почвы в дистиллированной воде в следующих пропорциях: для подстилки – 1:10 (первый эксперимент) и 1:50 (второй эксперимент), для почвы – 1:5 (первый эксперимент) и 1:10 (второй эксперимент). Растения культивировали в течение двух недель при постоянной температуре и искусственном освещении, ежедневно доливая воду в сосуды для поддержания начальных условий разведения. В каждый вариант изначально закладывали по 70 семян. В конце эксперимента у каждого проростка измеряли длину корня с точностью до 1 мм, а также отмечали наличие некрозов. Объем выборки при биотестировании подстилки в первом эксперименте составил 2330 проростков, во втором – 4125 (для почвы 4425 и 2500 соответственно).
Зависимость между загрязненностью подстилки и реакцией биотеста в масштабе всего градиента хорошо описывается S–образной логистической кривой (Рисунок 19). В пределах буферной зоны максимальные значения токсичности соизмеримы со значениями в импактной зоне; в свою очередь, минимальные значения равны фоновому уровню. При оперировании индексом нагрузки, который рассчитан по концентрациям подвижных форм металлов обнаруживаются “выпадающие” образцы из импактной зоны с низкой фитотоксичностью, которая соответствует высокому содержанию металлов. Однако именно в данных образцах зарегистрированы очень низкие для импактной зоны значения кислотности: рН равен 5.5, в то время как для остальных – он лежит в интервале 3.7 – 4.5. Использование индекса нагрузки, рассчитанного по обменным формам металлов, все расставляет по своим местам: прежде выпадающие точки из импактной зоны сдвигаются к облаку буферных точек и логистическая функция значительно лучше аппроксимирует дозовую зависимость .
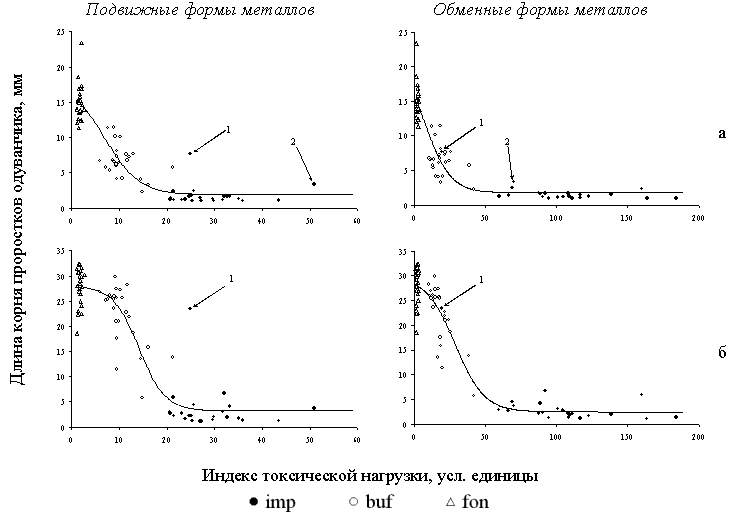
Рисунок 19. Зависимость результатов корневого теста от содержания тяжелых металлов (индекс токсической нагрузки) при тестировании образцов лесной подстилки из фоновой (fon), буферной (buf) и импактной (imp) зон при разной концентрации суспензии: а – отношение подстилки к воде равно 1:10, б – 1:50. Линия – аппроксимация зависимости логистическим уравнением регрессии. Стрелками показаны “выпадающие” образцы из импактной зоны.
Результаты биотестирования почвы существенно отличаются от результатов для подстилки. Различия касаются двух аспектов: очень большого разброса токсичности в фоновой зоне и крайне нестабильного “положения” образцов из буферной зоны при разной концентрации суспензии. При выяснении причин большого разброса между образцами фоновой зоны оказалось, что аномально низкие значения длины корня, равные значениям при биотестировании почвы из импактной зоны, связаны не с различиями в концентрациях тяжелых металлов, а с высокими значениями гидролитической кислотности. Учитывая тесную связь разных параметров почвенного поглощающего комплекса между собой, можно предположить, что разброс в фоновых образцах связан с известным эффектом эндогенной токсичности почвы, обусловленным повышенным выходом в почвенный раствор алюминия и марганца при увеличении кислотности. Неустойчивое “положение” в дозовой зависимости образцов из буферной зоны важно в контексте обсуждаемой проблемы триггерного эффекта. При менее концентрированной суспензии облако буферных точек полностью совпадает с облаком фоновых, а при более концентрированной – импактных. Следовательно, относительно небольшое смещение параметров почвенного раствора “переключает” реакцию биотеста с фонового уровня на импактный.
Безусловно, использованный нами корневой тест – очень упрощенная модель реакции биоты на токсическую нагрузку. Тем не менее, в полученной дозовой зависимости наблюдается выраженный триггерный эффект: в буферной зоне разброс фитотоксичности покрывает почти весь диапазон возможных значений, а переход от фонового уровня к импактному занимает небольшую долю от общей величины всего градиента нагрузки. Следовательно, пространственное варьирование концентраций поллютантов в буферной зоне даже в пределах относительно небольшого участка приводит к возникновению нелинейности в реакции биоты и может служить объяснением триггерного эффекта. Учитывая, что при биотестировании использована генетически однородная выборка, можно говорить о том, что именно высокое пространственное варьирование содержания токсичных форм тяжелых металлов, которое, в свою очередь, определяется наложением двух пространственных мозаик – валового содержания и кислотности субстрата – ключевой механизм возникновения триггерного эффекта в реакции биоты на токсическую нагрузку.
| Дальше
|
Назад |
Начало |
Список
|