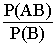 =
= 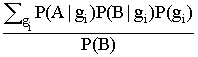 .
.Опубликовано: Основные положения диссертации на соискание ученой степени доктора биологических наук по специальности 03.00.16 – экология, 2004 г.
БИОХОРОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ И МОДЕЛИРОВАНИЕ ПРОСТРАНСТВЕННОЙ СТРУКТУРЫ РАСТИТЕЛЬНОГО ПОКРОВА (ГЕОИНФОРМАЦИОННЫЙ ПОДХОД)
Савельев А.А.
( Кафедра моделирования экологических систем экологического факультета Казанского государственного университета )
СОДЕРЖАНИЕ
Общая характеристика работы
1. Описание, анализ и моделирование структуры и хорологии природных систем
2. Ландшафтно-экологическая характеристика территории исследования. Исходные данные и их формализация
3. Вероятностная модель флористического состава растительного покрова
4. Градиентный анализ видов флоры и ординация растительности
5. Отклик видов флоры на факторы среды и его моделирование на исследуемой территории
6. Моделирование пространственного распределения видов флоры сосудистых растений
Выводы.
Список основных публикаций
Общая характеристика работы
Актуальность темы. Глобальная проблема сохранения биоразнообразия (в рамках видов, между видами и разнообразия экосистем) была осознана человечеством в прошлом веке, что привело к выработке международным сообществом Конвенции о Биологическом Разнообразии и разработке национальных стратегий изучения и охраны флоры и фауны отдельных территорий. Все возрастающее антропогенное давление на природу разрушает исторически сложившиеся ландшафтно-экологические закономерности, ведет к исчезновению видов, уничтожению и дигрессивному разрушению природных экосистем. Дальнейшее сокращение биоразнообразия может привести к дестабилизации биологических сообществ, утрате способности биоты поддерживать важнейшие функции биосферы и ее целостность. В результате необратимого перехода биосферы в новое состояние условия жизни на планете могут оказаться непригодными для человека.
Все вышесказанное относится и к сосудистым растениям, которые играют важнейшую экологическую роль в биосфере с точки зрения существования и устойчивости наземных экосистем на всех уровнях, от топологического до глобального, что делает несомненно актуальной задачу изучения, оценки и моделирования их пространственного распространения. Ее успешное решение требует не только сбора и накопления значительного по объему фактологического материала, но и его осмысления с целью выявления региональных и экотопологических закономерностей структуры растительного покрова, теоретико-методической проработки вопросов обеспечения корректного пространственного анализа и моделирования.
Возросшая в последнее время доступность и полнота данных об окружающей среде, в первую очередь данных дистанционного зондирования Земли (ДДЗЗ), создание компьютерных баз данных о растительности (базы данных геоботанических описаний, типов растительных сообществ, пространственных единиц растительности), а также современные вычислительные и информационные технологии открывают новые возможности для решения проблем сохранения биоразнообразия и контроля состояния экосистем на всех уровнях. Это касается и возможностей построения оценочных и прогностических пространственных моделей растительности с детальностью до отдельных видов на основе математического аппарата пространственного анализа и моделирования.
Нами представлены результаты исследований в области пространственного анализа и моделирования геосистем в рамках основного научного направления экологического факультета Казанского государственного университета по теме “Пространственно-функциональная структура флоры и растительности фрагментированных ландшафтов зонального экотона”, регистрационный номер 01.200.120118. Отдельные аспекты исследований поддержаны грантами Российского Фонда Фундаментальных Исследований (РФФИ) 1995-1997 гг. по теме: "Эрозия почв на востоке Русской равнины" (№ 95-05-153802), РФФИ 2003-2004гг. по теме: "Динамика пространственной структуры биоразнообразия экологических регионов РТ" (№ 03-04-96235), и Всемирного фонда дикой природы 2003г. по теме “West Pacific Gray Whales conservation, Sachalin, Russia” (№ M3979/RU008203/GLP).
Цель и задачи исследования. Целью работы явилось создание новых количественных моделей и методов для анализа и моделирования флористических данных на основе представлений о континуальности видового состава растительного покрова и стохастическом характере его пространственного распределения.
Для достижения поставленной цели необходимо было решить следующие задачи:
Исходные материалы и методика исследований. Выбор территории Республики Татарстан (РТ), расположенной в пределах зонального экотона, в качестве модельной анализируемой, обусловлен выраженной дифференциацией ландшафтного разнообразия с наличием плавных изменений состава растительности и факторов среды природных комплексов, представляющих зональные фрагменты хвойных, хвойно-широколиственных, широколиственных лесов и лесостепи, что позволяет показать особенности предлагаемой вероятностной модели. Исходные данные были пространственно унифицированы и интегрированы автором в единую геоинформационную базу данных
В качестве фактологической основы работы использовались базы данных и электронные карты, созданные на Экологическом факультете КГУ при активном участии автора:
Кроме того, использовались цифровая модель рельефа, полученная в результате топографической миссии космического челнока (совместный проект NASA и NIMA, США), и данные дистанционного зондирования Земли с приборов TM и ETM+ (спутники Landsat) и MODIS (спутники EOS Terra), предоставленные ИТЦ СканЭкс (Москва).
В работе широко использовались разнообразные методы исследования: эколого-флористический, картографический, математического моделирования, методы математической статистики, многомерной классификации и пространственного анализа (в том числе с применением искусственных нейронных сетей). Обработка данных проводилась с использованием лицензионных программ “MS Access”, "SURFER 7", "MapInfo Professional 6.0", "STATGRAPHICS Plus", CANOCO, BioMapper и программного обеспечения, созданного с нашим участием.
Научная новизна и теоретическая значимость заключаются в решении крупной научной проблемы фундаментальной экологии, а именно в выявлении закономерностей формирования биохорологического разнообразия и создании комплексного подхода к моделированию растительного покрова и прогнозу его пространственной структуры в условиях зонального экотона.
Впервые на основе гипотезы об экологической индивидуальности видов сформулирована вероятностная модель флористического состава растительного покрова как совокупности индивидуальных стохастических откликов видов на экологические условия среды. Показана адекватность этой гипотезы фактическим данным, полученным на модельной территории.
На основе вероятностной модели разработаны методы моделирования флористического состава растительного покрова, базирующиеся на комплексном использовании методов оценки отклика видов и учета пространственной стохастической составляющей их распределения.
Впервые предложен метод непрямого градиентного анализа растительных сообществ по флористическому составу, основанный на созданной вероятностной модели и методах построения топографических отображений видового состава на ординационную плоскость. Метод совмещает математическую классификацию и ординацию сообществ и позволяет отобразить континуальность изменения флористического состава.
Впервые для пространственного моделирования флористического состава растительного покрова на единой методической основе разработан и реализован в виде комплекса программ широкий спектр математических моделей, совмещающих традиционные подходы к моделированию отклика видов на факторы среды с геостатистическими методами учета типичной пространственной мозаики распределения видов и моделирования пространственной неопределенности.
Впервые показана возможность использования математического аппарата топографических отображений для автоматического моделирования и пространственной оценки видового состава растительного покрова непосредственно по данным ДДЗЗ без пересчета их в факторы среды.
Впервые реализован в геоинформационной базе данных алгоритм пространственной оценки полного видового состава растительного покрова (актуального пула видов), и выполнена такая оценка на модельную территорию.
Основные научные результаты исследований:
Практическая значимость работы. Разработанные методы использовались для комплексной экологической оценки и районирования территории в рамках проекта "Количественная оценка воздействия на окружающую среду объектов ОАО "Татнефть" методами геоинформационных технологий" (1995 – 1998 гг.). Методы автоматизированной дешифровки ДДЗЗ на основе нейронные сетей Кохонена, реализованные в программе Scanex NeRIS, применялись для построения карты малонарушенных лесов масштаба 1:1 000 000 на территорию всей России (Aksenov et al., Atlas of Russia’s Intact Forest Landscapes, Москва: 2002, 186 стр.), использовались в подразделениях МПР РФ при реализации “Федеральной целевой программы развития минерально-сырьевой базы РФ на 1994-2000 г.г.” (договора 120-00 от 30 марта 2000 г. и 0025-01 от 22 ноября 2000 г.), были рекомендованы как типовые для решения ряда задач в “Классификаторе тематических задач оценки природных ресурсов и окружающей среды, решаемых с использованием материалов дистанционного зондирования”, утвержденном МПР РФ в 2001г.
1. Описание, анализ и моделирование структуры и хорологии природных систем
Современные подходы к описанию природных систем, связанные с анализом видовового разнообразия растительного покрова, основаны, в первую очередь, на методах моделирования пространственных данных. Поскольку решающую роль в формировании видового состава растительного покрова на региональном и локальном уровнях играют факторы среды, основное внимание уделяется формализации описания такой связи и ее использованию для пространственного моделирования.
В научной среде географов и экологов существуют различные подходы как к методологии описания пространственных аспектов различных природных систем, так и к самому разделению экологических и географических аспектов такого описания (представление о биогеоценозах как элементарных биохорологических единицах биосферы, основанное на работах В.И.Вернадского, В.В.Докучаева, В.Н.Сукачева, Н.В.Тимофеев-Ресовского; экосистемный подход, известным представителем и популяризатором которого является Одум; учение о геосистемах Сочавы; ландшафтный подход, развитый в работах А.Г.Исаченко).
Подход к описанию растительности и ее разнообразия развит в работах Уиттакера и работах отечественных авторов Б.А. Юрцева и Л.И.Малышева. Описание связи факторов среды и растительности основывается на предложенном Уиттакером изучении комплексных градиентов среды как определяющих структуру сообществ, определении характеристик местообитаний и формализации понятия экологической ниши вида, введенного в работах Гринелла, Элтона, Гаузе и Хатчинсона. Для распределения растительности по градиентам среды рассматриваются две концепции: континуальности (основанная на гипотезе экологической индивидуальности видов и представленная в работах Л.Г.Раменского и Глизона), и холистическая, представленная в работах Клементса и Добенмайра.
Пространственная структура растительного покрова анализируется как на популяционно-видовом уровне, связанном с ординацией (работы Л.Г.Раменского, Б.М.Миркина, Г.С.Розенберга, Уиттакера и Кертиса), так и на фитоценотическом уровне, связанном с классификацией, различные подходы к которой представленный в работах Браун-Бланке, Добенмайра, В.Н.Сукачева, Б.М.Миркина, Т.А.Работнов, Т.И.Исаченко.
В настоящее время активно развиваются методы градиентного анализа сообществ (неформальные методы, методы, основанные на расстоянии, непрямой и прямой градиентный анализ, развитые в работах Уиттакера, Кертиса, Орлочи, Гауча, Хилла, Тер Браака), позволяющие численно описать связь факторов среды и отклика на них растительности. Возможен и надфитоценотический уровень хорологических единиц структуры растительности, связанный с анализом пространственной мозаики, и вопросами картирования.
Все это приводит к существующим концептуальным подходам к моделированию пространственной структуры растительного покрова как отклика на градиенты факторов среды, включая равновесные и неравновесные модели, индивидуалистский и холистический подходы, использование концепции ниш, способы моделирования местообитаний и пространственной мозаики на различных масштабных уровнях, и выбор предикторов пространственного распределения видов и единиц растительности.
Когда модель отклика отдельных видов или сообществ на факторы среды построена, традиционным способом моделирование на ее основе потенциального пространственного распределения видов или сообществ является моделирование их потенциальных местообитаний с последующим переходом к растительности. Показана ограниченность такого подхода – так, традиционные методы моделирования пространственного распределения растительности являются детерминированными, и позволяют воспроизвести только усредненное пространственное распределение. Очевидно, что при этом возникает проблема утери специфической стохастической структуры растительного покрова, проявляющейся на наиболее детальных уровнях масштабной иерархии, и связанной со случайными причинами, которые приводят к появлению характерной пространственной мозаики; существующие отдельные работы не позволяют полностью решить эту проблему.
Таким образом, необходима разработка комплексного подхода, позволяющего описывать растительность с использованием формальных единиц заданной детальности, с учетом связи этих единиц с факторами среды, с возможностью описания и моделирования реально существующей на конкретной территории стохастической пространственной мозаики растительного покрова в различных масштабах, и совмещающего достоинства холистического и индивидуалистского подходов к моделированию растительности
2. Ландшафтно-экологическая характеристика территории исследования. Исходные данные и их формализация
Исследования проводились на двух общепринятых уровнях пространственной детализации, что соответственно определило два варианта масштабов исследования: 1) региональный уровень (М 1:200 000) — территория в административных границах Республики Татарстан;
2) топологический (локальный) уровень (М 1:50 000-1:25 000 и крупнее) — территория Раифского участка Волжско-Камского государственного природного заповедника.
Приведем основные ландшафтно-экологическая характеристики территории исследования для регионального и локального уровня, способы представления пространственной информации в геоинформационных системах и методы пространственной унификации данных, описание данных о растительном покрове.
Основными источниками информации о флоре и растительности РТ явились авторские флористические и геоботанические описания, гербарные сборы, гербарные коллекции, хранящиеся в Гербариях КГУ, МГУ, Главного Ботанического сада (Москва), БИН РАН (С-Петербург), Казанского государственного педагогического университета, Елабужского государственного педагогического института, Ижевского государственного университета и др.; публикации, выходившие в свет в период с середины прошлого века до 1997-98 гг., и посвященные результатам разномасштабных обследований территории и отдельным находкам видов сосудистых растений в границах РТ. В результате обработки информации описаний и находок для территории республики выявлено 1610 видов природной и натурализовавшейся флоры сосудистых растений, а также 80 видов, наиболее часто возделываемых человеком и способных в перспективе пополнить состав природной флоры. Современный конспект видов содержится в последней флористической сводке О.В. Бакина, Т.В. Роговой, А.П. Ситникова “Сосудистые растения РТ”. На основе этих данных на экологическом факультете КГУ была создана геоинформационная флористическая база данных (БД) “Флора”, включающая характеристики 3160 площадок описаний, перечень видов на них и справочник по видам флоры. Всего описано 1690 видов флоры. Латинские названия видов, также как и латинские названия родов и семейств, даются по сводке С.К.Черепанова "Vascular Plants of Russia and Adjacent States (the forms USSR)".
Численное представление в единой геоинформационной базе данных характеристик окружающей среды для регионального – РТ, и локального – Раифа, масштабов, включая обработку данных дистанционного зондирования Земли (геометрическая коррекция, расчет температуры и вегетационных индексов), выполнено автором (указаны характеристики, использованные для расчетов в каждом масштабе).
Перечень использованных в работе характеристик окружающей среды дан в таблице 2.1.
Таблица 2.1.
|
РТ |
Факторы среды |
Раифа |
|
X1 |
температура поверхности Земли (градус К) - весна |
|
|
X2 |
температура поверхности Земли (градус К) – лето |
X1 |
|
X3 |
температура поверхности Земли (градус К) – осень |
X2 |
|
индекс влажности - весна |
||
|
индекс влажности - лето |
X3 |
|
|
индекс влажности - осень |
X4 |
|
|
индекс “зелености” - весна |
||
|
индекс “зелености” - лето |
X5 |
|
|
индекс “зелености” - осень |
X6 |
|
|
X4 |
нормализованный разностный вегетационный индекс - весна |
|
|
X5 |
нормализованный разностный вегетационный индекс - лето |
X7 |
|
X6 |
нормализованный разностный вегетационный индекс - осень |
X8 |
|
X7 |
годовая суммарная радиация (мДж/м2) |
|
|
X8 |
сумма биологически активных температур (0С) |
|
|
X9 |
сумма осадков в теплый период года (мм) |
|
|
X10 |
модуль поверхностного стока (мм/год) |
|
|
X11 |
максимальная высота снежного покрова (см) |
|
|
X12 |
валовое увлажнение территории (мм/год) |
|
|
X13 |
коэффициент стока |
|
|
X14 |
гидротермический коэффициент |
|
|
X15 |
абсолютная высота (м) |
X9 |
|
X16 |
экспозиция склона |
X10 |
|
X17 |
крутизна склона (градус) |
X11 |
|
X18 |
тип местности |
|
|
X19 |
тип почв |
|
|
X20 |
гранулометрический состав почв (ГМС) |
|
|
X21 |
преобладающий состав подстилающих горных пород |
|
|
X22 |
мощность четвертичных отложений (м) |
3. Вероятностная модель флористического состава растительного покрова
Главным объектом изучения и моделирования является флористический состав растительных сообществ, представленных на пространственных площадках, характерный размер которых соответствует описаниям БД “Флора”. Важность изучения именно флористического состава растительности неоднократно обосновывалась различными авторами, в первую очередь потому, что он связан с биоразнообразием, и моделирование пространственного распределения видов является важной задачей в рамках мер по его сохранению. Другим аргументом в пользу выбранного подхода является большая устойчивость и объективность флористического состава по сравнению с различных оценок обилия видов (Василевич 1976, 1992; Юрцев, Семкин 1980; Пааль, Семкин 1983; Малышев 1975, 1992)..
Модель флористического состава построена в рамках следующих предположений, которые подтверждаются анализом данных, имеющихся в БД “Флора”:
Поскольку в соответствии с концепцией континуума в условиях гетерогенной абиотической среды отмечается стохастическое распределение видов по локальным градиентам факторов согласно их экологическим потребностям (Раменский, 1938; Уиттекер, 1980), равновесное состояние предполагается не детерминированным, а стохастическим, т.е. случайные отклонение от равновесия имеют пространственно распределенную случайную составляющую, что позволяет использовать для пространственного анализа и моделирования методы геостатистики.
Начиная с 1980 г. в зарубежной биогеографической литературе для характеристики флористического состава отдельных участков земной поверхности стал использоваться термин "актуальный пул видов - actual species pool" (Abbott, 1977; Abbott, Black, 1980). Согласно теории пула видов, реально наблюдаемое в местообитании флористическое разнообразие, зафиксированное в конкретном описании пробной площади, может и должно быть дополнено видами, способными существовать в данных экологических условиях, и отмечаемыми на смежных территориях. Воспроизводимая совокупность видов представляет локальный пул видов (E.Van der Maarel, 1997; J.Ewald, 2002).
Поскольку даже для одной точки территории списки видов, отмеченных в описаниях разных авторов, либо сделанных в разное время, обычно различаются, эти описания можно рассматривать как выборку из генеральной совокупности всех возможных сочетаний видов локального актуального пула, распределение вероятностей которой определено сочетанием экологических факторов в данной точке.
Таким образом, важной является задача оценки видового состава актуального пула по отдельным описаниям. Учитывая, что для каждой точки территории имеются обычно только одиночные описания, сделать это можно лишь используя информацию о структуре регионального пула видов, содержащуюся во всей совокупности описаний для данной территории.
Для описания видового состава растительного покрова обычно используются флористические списки видов. Будучи дискретными, они не всегда удобны для анализа и моделирования в силу континуального характера изменения растительности, поэтому часто используются их вероятностные аналоги. Способы получения вероятностного описания флористического состава растительного покрова известны: для анализа и классификации это сглаживание по Beals (Beals, 1984), для прогнозного моделирования – логистическая регрессия и другие аналогичные модели, использующие данные о значениях факторов среды на площадках.
Построение вероятностной модели в рамках гипотезы индивидуальности видов
Предложена модель вероятностного описания флористического состава растительного покрова, которая, при выполнении гипотезы индивидуальности, для получения списка вероятностей присутствия каждого вида на площадке не требует данных о факторах среды, а использует только сами флористические списки.
Модель формулируется следующим образом.
Пусть g Î G - некоторое значение комплексного фактора среды (сочетание значений всех факторов среды на данной площадке). Вероятность наличия вида А в условиях g обозначим как Р(А|g) или Pg(A).
Предположим, что справедлива гипотеза индивидуальности:
1) Вероятность присутствия вида на площадке не зависит от присутствия или отсутствия на ней других видов (при этом рассматриваются только парные зависимости между видами, поскольку в статистическом анализе они традиционно предполагаются наиболее значимыми): Pg(A)=Pg(A|B),
2) Вероятность наблюдать тот или иной вид на данной площадке зависит только от условий среды g на ней.
Выразим вероятность Р(А|g) через условные вероятности P(A|B), связывающие присутствие разных видов. Условная вероятность Р(А|В) наблюдать вид А при наличии вида В для всей территории есть:
Р(А|В) = = .
Используя формулу Байеса, перепишем уравнение условной вероятности:
|
P(A|B) = |
(3.1) |
Обозначим список видов, присутствующих на некоторой площадке с условиями среды g, как Sg. Тогда вероятность присутствия вида А на площадке можно оценить как
|
Р(А|g) |
(3.2) |
Это следует из выражения (3.1), поскольку условная вероятность P(A|B) для фиксированного A будет максимальна при таком виде B, который является индикаторным видом для оптимальных условий среды вида A. Для такого B вероятность P(gi|B) будет близка к единице для некоторого i (поскольку сумма этих вероятностей по всем gi равна единице), причем условия среды gi почти всегда будут совпадать с условиями среды g анализируемой площадки. Следовательно, в сумме только одно слагаемое при gi=g будет существенно больше нуля, и сама сумма будет примерно равна Р(А|g).
Сами вероятности P(A|B) для каждой пары видов A и B могут быть легко оценены на основе флористических списков с использованием частоты: они вычисляются как отношение числа случаев совместного нахождения видов А и В к числу всех случаев нахождения вида В. Далее, используя формулу (3.2), можно перейти от флористических списков к вероятностным спискам видов, заменяя отсутствующие в списке виды оценкой вероятности Р(А|g) их присутствия.
Проверить адекватность предложенной модели можно проверкой справедливости основного утверждения, т.е. Pg(A)=Pg(A|B). Напрямую это сделать затруднительно, так как надежная оценка этих вероятностей требует многократных повторных наблюдений для всех видов в конкретных условиях среды, и для проверки лучше использовать величины, рассчитываемые по всей совокупности данных.
Условную вероятность наблюдать вид А при наличии вида В для данного распределения условий среды G по территории и выполнении гипотезы индивидуальности, вычисленную согласно приведенной выше формуле (3.2) будем называть модельной вероятностью.
Имея только данные о присутствии/отсутствии видов на площадках, для каждой пары видов A и B можно оценить вероятность Р(А|В) напрямую с использованием частоты, будем называть вероятность, оцененную таким способом, фактической вероятностью.
Фактические и модельные вероятности были оценены нами на реальных данных, взятых из базы “Флора” и показали хорошее согласование. Факторы среды оценивались для каждой площадки по шкалам Цыганова, для статистических вычислений площадки объединялись в 37 и 71 класс по сходству факторов среды методом иерархической кластеризации Варда.
Адекватность предложенной модели имеющимся фактическим данным оценивалась путем вычисления коэффициента корреляции между модельной и фактической вероятностями по всем парам видов. Кроме того, оценивалась применимость данной модели для видов с различной распространенностью. Для этого коэффициент корреляции между модельной и фактической вероятностью был вычислен при различных пороговых значениях встречаемости; результаты представлены в таблице 3.1. Высокие значения коэффициентов корреляции свидетельствуют в пользу адекватности модели.
Таблица 3.1.
|
Оценка коэффициента корреляции модельной и фактической вероятностей в зависимости от встречаемости видов. |
||||
|
37 Классов |
71 Класс |
|||
|
Число площадок, на которых встречены виды |
Коэф. корреляции R |
Число пар |
Коэф. корреляции R |
Число пар |
|
1 и более |
0.69 |
722436 |
0.71 |
657604 |
|
2 и более |
0.75 |
663268 |
0.77 |
612772 |
|
3 и более |
0.79 |
608994 |
0.80 |
569140 |
|
4 и более |
0.82 |
530016 |
0.83 |
501820 |
|
7 и более |
0.86 |
444788 |
0.87 |
426952 |
|
12 и более |
0.89 |
351154 |
0.90 |
340690 |
|
20 и более |
0.91 |
249072 |
0.92 |
244886 |
|
33 и более |
0.93 |
165082 |
0.94 |
163462 |
|
55 и более |
0.94 |
109380 |
0.95 |
108986 |
|
90 и более |
0.95 |
61750 |
0.96 |
61732 |
|
150 и более |
0.96 |
30800 |
0.96 |
30800 |
|
245 и более |
0.96 |
12882 |
0.97 |
12882 |
|
400 и более |
0.97 |
3306 |
0.97 |
3306 |
|
550 и более |
0.95 |
870 |
0.95 |
870 |
|
660 и более |
0.92 |
420 |
0.92 |
420 |
|
810 и более |
0.89 |
110 |
0.89 |
110 |
Применение вероятностной модели
Предлагаемая модель позволяет, за счет использования статистических суррогатов (вероятностных списков видов), аппроксимировать континуальное распределение растительности на территории, что снижает влияние случайных изменений в видовом составе и повышает статистическую устойчивость получаемых результатов.
Можно охарактеризовать следующие основные пути практического применения построенной модели:
1. Вопрос классификации растительности является одним из сложнейших, и выполнение классификации представляет собой отдельную трудную задачу. Предложенная модель позволяет не выделять классы сообществ, а моделировать пространственное распределения различных видов независимо.
2. Предложенная модель дает обоснованный метод (формула 3.2) для перехода от флористических списков к вероятностным, которые лучше отражают континуальность видового состава растительного покрова и позволяют уменьшить влияние так называемой “проблемы двойных нулей” при анализе видового состава и его связи с факторами среды. Проблема, как известно, состоит в том, что, не имея дополнительной информации, мы не можем установить, чем вызвано отсутствие данного вида на данной площадке: неблагоприятными условиями среды, в которых произрастание данного вида невозможно, либо случайными факторами при благоприятных условиях. Отсутствие такой информации не позволяет полно оценить сходство видового состава растительности на разных площадках, и существенно искажает результаты анализа.
Сравнение результатов ординации видов, представленных в базе данных “Флора”, методом анализа соответствия (Legendre и Legendre, 1998) по флористическим и вероятностным спискам показало, что использование вероятностных списков позволяет значительно снизить общую вариацию данных и получить лучшую интерпретацию результатов за счет меньшего числа ординационных осей, необходимых для их представления.
3. Модель (формула 3.2) использовалась для прогноза вероятности присутствия на площадках видов, информация о которых отсутствует (например, они не определялись, и для них нет данных ни о присутствии, ни об отсутствии), по тем видам, которые на данной площадке обнаружены. Сравнение результатов прогноза с другими моделями, основанными на моделировании отклика видов на факторы среды, показало ее высокие прогностические свойства.
4. Градиентный анализ видов флоры и ординация растительности
С последней четверти прошлого века развиваются методы непрямого градиентного анализа (метод главных компонент, анализ соответствий, факторный анализ, метод Орлоци, метод Гудола, Висконсинская и Гауссова ординация (Розенберг, Миркин, 1978; Миркин, 1985; Legendre,1998)), а затем и прямого градиентного анализа (различные виды регрессии, канонический анализ соотвествия, (ter Braak, 1986, Legendre,1998)), которые позволили работать с нелинейными зависимостями в многомерных данных, и ставшие в наше время стандартным инструментом изучения растительности.
Для изучения растительного покрова впервые для территории Республики Татарстан был проведен градиентный анализ видов флоры сосудистых растений из БД “Флора”. При проведении анализа были рассмотрены два способа представления флористических данных – флористические списки и вероятностные списки, и два ординационных метода - анализ соответствий и канонический анализ соответствий, в результате чего получены четыре варианта ординации видов. Для реализации методов использовалась программа Canoco (Leps, Smilauer; 2003) и средства языка R (Ripley; 2001).
Структура результатов ординации видов для непрямого градиентного анализа оказалась похожей, а сама ординация достаточно выраженной как для флористических, так и для вероятностных списков - собрались в отдельные кластеры лесные виды, водные и болотные виды, рудеральные и культурные виды, луговые виды, степные виды; эколого-ценотические группы плавно переходят друг в друга. В то же время, в силу формальности примененных алгебраических методов, ординационные оси для двух вариантов представления данных оказались развернутыми друг относительно друга, а возможная интерпретация осей в обоих случаях – неочевидной и различной.
Так, для флористических списков первая ось ординации интерпретируется как некий комплексный градиент, проходящий от бореальных через суббореальные к аридным местообитаниям. Вдоль второй оси ординации виды расположились в следующей последовательности: виды каменистых степей ® степные виды® лугово-степные виды® луговые виды® лесолуговые и лесные виды® виды влажных лугов® болотные и водные виды, что позволяет интерпретировать ее как градиент влажности или увлажнения, проходящий от наиболее сухих к наиболее влажным местообитаниям. Для вероятностных списков первая ось ординации интерпретируется как градиент увлажнения. Расположение видов вдоль третьей оси ординации от степных и водных через луговые и лесные к рудеральным и культурным позволяет интерпретировать эту ось можно как градиент, характеризующий стабильность субстрата и проходящий от наиболее к наименее стабильным местообитаниям.
Больший интерес представляют для нас результаты прямого градиентного анализа, позволяющего установить и оценить связь видового состава и градиентов факторов среды. Для флористических списков первая ось ординации интерпретируется как зональный градиент, проходящий от более северных к более южным местообитаниям, и отражающий не только почвенные условия местообитания, но и климатическую составляющую – наибольший вклад здесь имеют тип почв, весенняя (майская) температура, гранулометрический состав почв, преобладающие породы и гидротермический коэффициент. Вторая ординационная ось может интерпретироваться как градиент высоты, проходящий от более низменных к более возвышенным местообитаниям, поскольку наибольший вклад для нее вносят абсолютная высота, тип местности, уклон, локальный базис эрозии. Для вероятностных списков первую ось ординации снова интерпретируется как зональный комплексный градиент, проходящий от северных (влажных и холодных с бедными “легкими” почвами) к южным (сухим и теплым с богатыми “тяжелыми” почвами) местообитаниям, поскольку наибольший вклад здесь вносят майская температура, тип почв, гранулометрический состав почв, гидротермический коэффициент. Расположение видов вдоль второй оси ординации от водных и болотных через лесные бореальные, лесные неморальные и луговые к степным, позволяет сделать вывод, что эта ось может интерпретироваться также как и вторая ось в предыдущем случае, т.е. как градиент высоты.
Обобщенные результаты сравнения способов представления данных приведены в таблице 4.1.
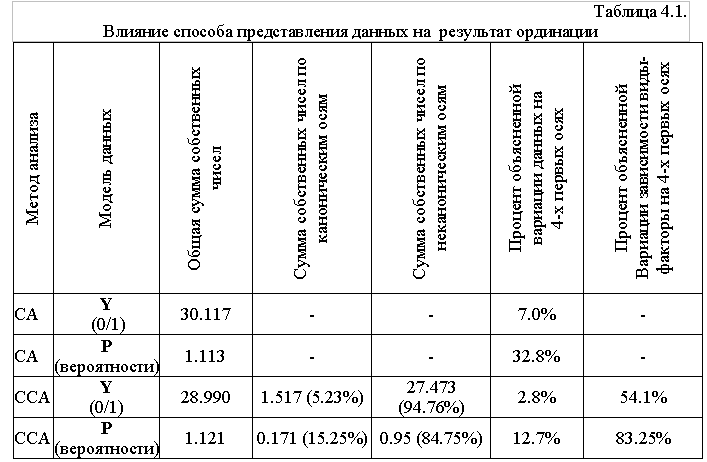
Сравнение сумм собственных чисел, отражающих общую изменчивость данных, позволяет сделать вывод о снижение их общей вариации при переходе к вероятностям. Так, для традиционно используемых первых четырех ординационных осей доля объясненной вариации для обоих методов анализа значительно выше при использовании вероятностных списков.
Непараметрическая классификация и ординация растительности
Поскольку одной из причин, приводящих к затруднению интерпретации ординационных осей, является использование методов линейной алгебры, был предложен непараметрический метод, совмещающий ординацию и классификацию сообществ, и позволяющий нелинейно представить все зависимости и градиенты на одной ординационной плоскости. При этом под термином “классификация” понимается разделение описаний на классы на основе сравнения их характеристик, без привлечения заранее определенных правил или систем классификации. Отметим, что при этом выделение единиц растительности, “адаптированных” к данной конкретной территории, не преследует целей построения классификации растительности или легенды карты, которая будет использована для представления информации человеку; одной из задач в нашем случае является сохранение и представление континуальности. В качестве исходных данных нами снова использовались как флористические, так и вероятностные списки.
Нейронные сети Кохонена как самоорганизующаяся модель выделения и ординации категорий
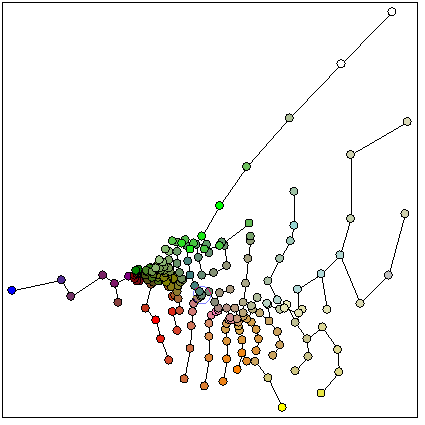
Для градиентного анализа был использован алгоритм “самоорганизующихся карт” (Self Organizing Map - SOM) Кохонена (Kohonen, 1997), относящийся к топографическим отображениям (под топографией здесь понимают сохранение порядка и взаимоположения по сходству объектов при отображении). Данный метод позволяет представить совокупность описаний в терминах абстрактных классов (суррогатных сообществ), представленных вероятностями видов и образующих регулярную решетку на ординационной плоскости.
Данный метод позволяет получить аппроксимацию континуального характера изменения флористического состава, а также отразить связь такого изменения с градиентами факторов среды. Для вычислений использовалось программное обеспечение, разработанное автором, и реализующее алгоритм SOM.
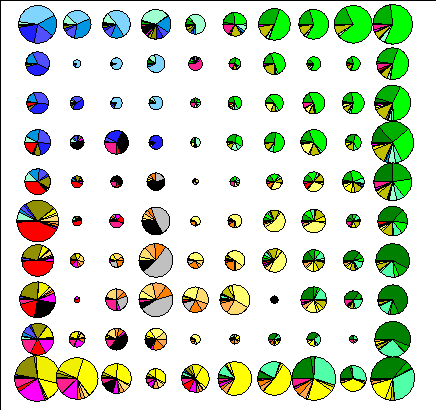
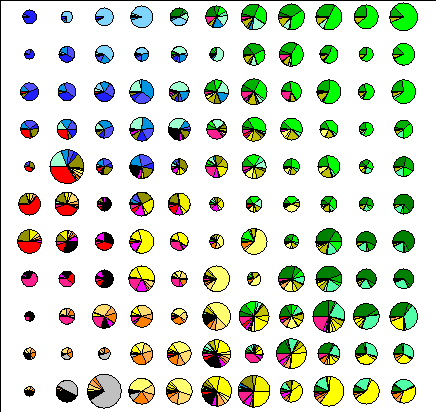
Размер решетки классов нейронной сети был выбран примерно одинаковым в обоих случаях (10x10 для флористических списков, и 11x11 для вероятностных списков). На рисунке 4.1 показана (круговыми диаграммами) эколого-ценотическая структура полученных классов и (размером диаграммы) общее количество площадок, отнесенных к каждому из них.
|
|
|
|
По флористическим спискам |
По вероятностным спискам |
|
Рис.4.1. Эколого-ценотическая структура классов. |
|
В соответствии с (Kohonen, 1997) размер классов в первом случай возрастает от центра к периферии, что является артефактом метода и говорит о смещении результатов в сторону классификации. Применение вероятностных списков позволяет получить равномерную аппроксимацию описаний классами, а баланс между ординацией и классификацией регулируется параметрами метода.
Нижний правый угол решетки полученного отображения занимают бореальные формации, причем доля сосняков в классах убывает снизу вверх. Далее, при продвижении вверх по ординационной плоскости, количество бореальных формаций в целом в классах убывает, но при этом доля ельников в общем количестве бореальных формаций увеличивается, а сосняков – уменьшается.
Правый верхний угол представлен неморальными сообществами. Здесь сосредоточены липняки, дубравы, осинники, встречаются кленовники. При продвижении вниз доля этих формаций в классах постепенно снижается. Если двигаться по ординационной плоскости от правого угла влево, можно наблюдать, что в классах увеличивается доля ольшанников, вязовников и ивняков, что соответствует градиентному ряду увеличения увлажненности.
Левая половина отображения представлена открытыми травянистыми формациями – степями, лугами, болотами. Верхняя левая часть состоит из классов, в которых наибольшую долю составляют низинные и сфагновые болота. Это область избыточного увлажнения. В классах этой группы встречаются низинные и пойменные луга, а также ивняки и березняки. Постепенно, при приближении к нижнему углу соотношение между болотами и низинными лугами увеличивается в пользу низинных лугов.
|
|
Бореальная эколого-ценотическая группа |
|
|
|
Ель финская (Picea fennica X.) |
|
|
|
Черника (Vaccinium myrtillus L.) |
|
|
|
|
|
|
По флористическим спискам |
По вероятностным спискам |
|
|
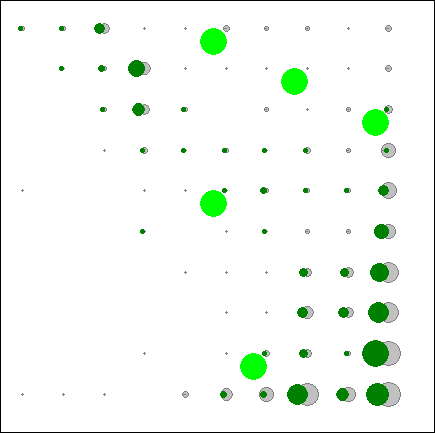
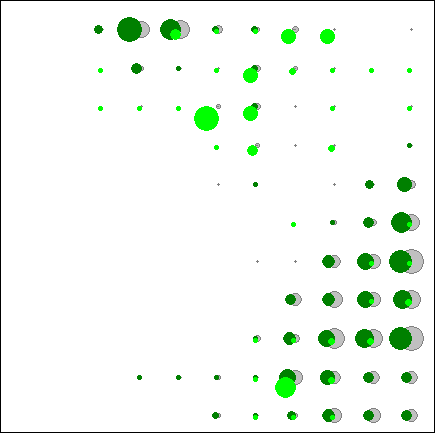
Рис.4.2. Распределение экспертно выбранных видов бореальной эколого-ценотической группы по классам нейронной сети. |
||
Таким образом, на отображении можно одновременно выделить несколько градиентных переходов, обусловленных факторами среды: по увеличению увлажнения (от степной группы через суходольные и низинные луга к болотам), по увеличению плодородия почв (от бореальной группы к неморальной), по уменьшению освещенности местообитаний (от лугов к лесам), по увеличению увлажнения (от неморальных лесов к болотам).
Этот вывод подтверждается и анализом распределения по классам нейронной сети отдельных видов, которые выбирались экспертно как наиболее типичные (на рис.4.2. приведено распределение двух видов бореальной ЭЦГ).
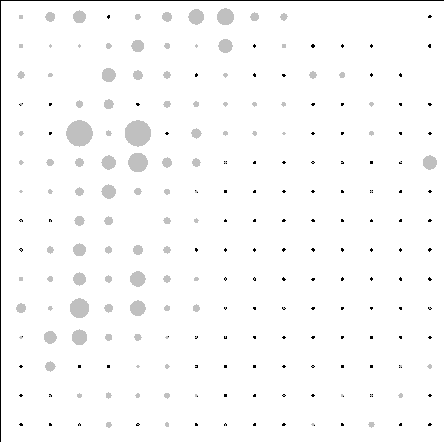
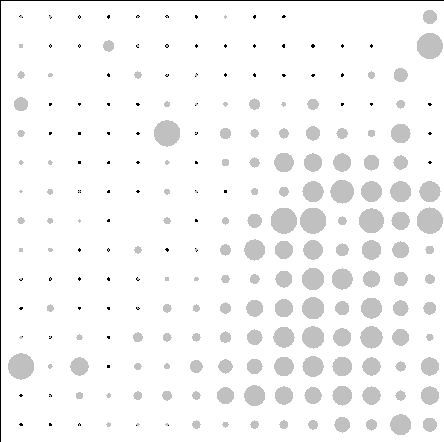
Полученную ординационную плоскость можно использовать и для отображения значения факторов среды для каждого класса, что позволяет отображать и анализировать связь классов (а следовательно и состава растительности) с градиентами факторов среды (рис.4.3).
Из сравнительного анализа распределений следует, например, что классам с преобладанием видов степных ЭЦГ соответствуют низкие значения гидротермического коэффициента, а с преобладанием видов бореальных ЭЦГ – высокие, что согласуется с использованием этого показателя для разделения территории по климатическим условиям на бореальную и лесостепную зоны. Для данной территории большие отметки высот соответствуют классам с преобладанием видов степных и неморальных ЭЦГ.
|
|
|
|
Гидротермический коэффициент |
Абсолютная высота |
Рис.4.3. Распределение значений факторов среды по классам нейронной сети.
Таким образом, предложенный метод с использованием SOM и вероятностных списков является развитием и обобщением существующих методов, и может быть использован при анализе растительности для получения аппроксимации континуального изменения флористического состава и оценки связи такого изменения с факторами среды.
Глава 5. Отклик видов флоры на факторы среды и его моделирование на исследуемой территории
В этой главе описаны статистические модели отклика видов флоры высших сосудистых растений на факторы среды. Построение моделей проводилось в локальном (топологическом) масштабе исследования четырьмя методами, и для каждого из них построены модели отклика для десяти выбранных видов флоры из различных ЭЦГ: белокрыльник болотный (Calla palustris L.), хамедафне прицветничковая (Chamaedaphne calyculata (L.) Moench.), ежа сборная (Dactylis glomerata L.), кислица обыкновенная (Oxalis acetosella L.), лютик кашубский (Ranunculus cassubicus L.), ольха серая (Alnus incana (L.) Moench.), пролесник многолетний (Mercurialis perennis L.), прострел раскрытый (Pulsatilla patens (L.) Mill.), страусник обыкновенный (Matteuccia struthiopteris (L.) Tod.), черника (Vaccinium myrtillus L.).
Модели оклика видов на факторы среды.
В качестве первой модели отклика использовалась логистическая регрессия (Dobson, 2002). Расчеты проводились с помощью программы Statgraphics Plus v.5.1 (Statgraphics…, 1995).
Анализ коэффициентов полученных моделей показал, что практически для всех видов значимы такие переменные как абсолютная высота, температура поверхности Земли летом и летний индекс влажности, а значения коэффициентов всех моделей согласуются с экологией видов. Так, если сравнить модели отклика черники и пролесника многолетнего, то можно заметить, что значения коэффициентов при переменных, соответствующих летней температуре и летней влажности, в этих двух моделях имеют противоположные знаки, что соответствует факторам, типичным для бореальных и неморальных сообществ соответственно. Для прострела раскрытого получены высокие положительные значения коэффициентов модели при летний температуры, летней влажности и экспозиции склона, что соответствует его приуроченности к теплым склонам.
В качестве второй модели использовались нейронные сети прямого распространения (Bishop, 1986), которые можно рассматривать, в том числе, и как усложнение логистической регрессии. Структура использованной нейронной сети включала 46 нейронов: 36 нейронов, объединенных в два слоя – 11 в первом и 15 - во втором, и 10 выходных нейронов, позволяющих одновременно моделировать отклики тех же десяти видов флоры, что и в случае логистической регрессии. Нейронная сеть обучалась методом обратного распространения ошибки в программе NP (Nevada Propogation, Goodman, 1996).
Так как нейронная сеть MLP реализует модель “черного ящика”, то интерпретация параметров обученной нейронной сети не представляется возможной.
В качестве третьей модели использовался факторный анализ экологических ниш, или ФАЭН (Perrin, 1984; Hausser, 1995; Hirzel, 2000), концептуальной основой которого является теория экологических ниш. Ниша вида описывается в терминах маргинальности (отклонения от средних значений факторов на данной территории) и специализации (за счет сопоставления разброса значений факторов по всей территории и в местах присутствия вида), которые поддаются интерпретации. Вычисления проводились с помощью программы Biomapper v.2.1 (Hirzel и др., 2002).
Так, для черники характерна выраженная отрицательная маргинальность по летним вегетационным индексам (причем с достаточно высокой специализацией), характеризующей фитоценозы с низкой летней и высокой осенней первичной продукцией по сравнению со средней для территории. Подобное распределение продукции по сезонам характернее для сообществ с доминированием хвойных лесообразующих пород, что соответствует экологии рассматриваемого вида. Аналогичный анализ для всех факторов показывает, что полученная модель экологической ниши черники соответствует большинству стандартных местообитаний черники на территории Раифы: склоны междюнных понижений в условиях эоловых форм рельефа песчаных террас в составе фитоценозов елово-сосновых зеленомошных лесов. Аналогичное соответствие было получено и для всех распространенных видов.
В то же время значения маргинальности и специализации для белокрыльника болотного не соответствуют ожидаемым для данного вида, поскольку модельная ниша слишком широкая, что может быть связано со статистическими погрешностями, обусловленными незначительной встречаемостью (5%) данного вида на площадках описаний исследуемой территории.
Для прямого использования данные ДДЗЗ, без определения на их основе факторов среды, был разработан новый метод, основанный на тематической калибровке генеративного топографического отображения классов яркостей снимка, полученного методом, являющийся развитием аппарата SOM (Generative Topographic Mapping или GTM, Bishop и др., 1998). Флористическая информация при построении отображения не используется, она нужна для его тематической калибровки: для этого используется признак присутствия конкретного вида на площадке, для которой известны яркостные характеристики снимка, а значит и положение на ординационной плоскости. Результатом калибровки является условная вероятность наличия вида на площадке, отнесенной по яркостным свойствам снимка к данному классу топографического отображения, т.е. искомая прогнозная модель наличия вида, представленная в виде непараметрической регрессии этой вероятности на яркостные свойства снимка территории.
Точность такой регрессии зависит, в частности, от точности представления яркостей классами, и растет с ростом числа классов и уменьшением ширины сглаживающего ядра. С другой стороны, генерализация полученной регрессии, то есть ее способность правильно отображать незнакомые данные, наоборот увеличивается с уменьшением числа классов и ростом ширины сглаживающего ядра. Выбор параметров построения отображения является важными для получения удовлетворительного компромисса между точность представления имеющихся данных и возможностью использования полученного топографического отображения для прогноза, и должен контролироваться экспертом. В качестве исходных данных для построения топографического отображения были совмещены летний и осенний снимки спутника Landsat (использовались 3, 4 и 5 каналы).
Значения яркостей были обработаны методом GTM, в результате чего каждому элементу изображения был присвоен номер класса топографического отображения на решетку размером 15x15=225 классов (рис.5.1).
|
|
|
|
Рис.5.1. Отображение Сэммона топографического отображения и результат классификации совмещенных снимков.
Вид отображения Сэммона достаточно типичен для покрытых растительностью территорий: выделяется наиболее спектрально темный класс, соответствующий чистой водной поверхности, от него идет “плотная” группа классов, соответствующих в основном хвойной лесной растительности, ниже нее – менее “плотная” группа классов, соответствующих другой растительности (в нашем случае – в основном лиственной, причем широколиственные и мелколиственные леса разделились), и на противоположной стороне отображения – наиболее спектрально яркие классы, соответствующие таким объектам, как жилая застройка, обнаженные выходы пород и т.д.
Для калибровки построенного топографического отображения по видам флоры использовались программы Scanex NeRIS и Timan, разработанных автором.
На рис.5.2 приведены результаты калибровки для четырех видов, относящихся к разным ЭЦГ: черника (лесной-бореальный), прострел раскрытый (лесной-боровой), пролесник многолетний (лесной-неморальный), белокрыльник болотный (водно-болотный). Размер значков пропорционален вероятности присутствия вида на ОТЕ, набор спектральных яркостей которой на снимках соответствует данному классу.
|
|
|
|
Черника |
Прострел раскрытый |
|
|
|
|
Пролесник многолетний |
Белокрыльник болотный |
Рис.5.2. Результаты калибровки топографического отображения.
Отметим, что топографические отображения обладают важным для анализа изображений свойством, а именно устойчивостью отображения по отношению к изменению фотометрических характеристик снимков, что позволяет использовать калиброванные отображения для анализа видового состава новых, но аналогичных использованным для калибровки, территорий, что делает такую модель пригодной для оперативной дистанционной оценки пространственного распределения видов.
Модель на основе топографического отображения может быть легко адаптирована для прогноза отклика не в терминах видов, а в терминах некоторых единиц растительности.
|
|
|
|
Бореальные сообщества (классы 7-10) |
Бореально-неморальные сообщества (классы 39-40) |
|
|
|
|
Неморальные сообщества (классы 99-100) |
Болотные сообщества (класс 91) |
Рис.5.3. Результаты калибровки топографического отображения классами SOM.
Для этого достаточно использования для калибровки другой тематической информации, связанной с площадками, например классов сообществ в некоторой системе классификации (единиц растительности) - например классов продромуса средней полосы России, классов ассоциаций, выделенных на основе доминантного подхода, или классы нейронной сети Кохонена, полученные при анализе флористического состава в главе 4 (рис.5.3); размер значков пропорционален вероятности. На рисунке приведены результаты калибровки для четырех групп классов нейронной сети Кохонена (SOM), которые содержательно интерпретировались в главе 4.
Приведем еще один пример применения метода, использовав для калибровки топографического отображения три типа ассоциаций Раифского участка ВКГПЗ: березняк осоково-снытево-пролесниковый, липняк осоково-пролесниково-снытевый, злаково-разнотравный луг, взятые с фрагмента крупномасштабной (1:10 000) карты растительности Раифского участка ВКГПЗ из фондов Экологического факультета КГУ.
В данном случае нами калибровалось топографические отображения, построенные для двух снимка по отдельности (рис.5.4.), что еще раз продемонстрировало устойчивость метода: для класса “березняк осоково-снытево-пролесниковый”, несмотря на существенные сезонные изменения, результаты калибровки летнего и осеннего снимков сходны (расположены приблизительно в одной области ординационной плоскости), хотя влияние сезонности приводит к “миграции” соответствующих классов по ней.
|
|
|
|
Березняк осоково-снытево-пролесниковый (осенний снимок) |
Березняк осоково-снытево-пролесниковый (летний снимок) |
Рис.5.4. Результаты калибровки топографических отображений для некоторых типов растительных ассоциаций.
Оценка адекватности и сравнение моделей
Для оценки качества моделей использовался тот же набор данных, что и для их калибровки. Для оценки качества строились таблицы сопряженности модельных данных с реальными с последующим вычислением общепринятых показателей (коэффициенты Симпсона, Рассела, Кульчинского, Брауна, Чекановского, Очаи, Сокала - для оценки правильности прогноза присутствия вида; Каппа, условная Каппа, коэфициенты Чупрова Юла, Роджерса, Хаммана, Микаэла и простой коэффициент связи – для оценки правильности прогноза как присутствия, так и отсутствия вида (Cox,Cox; 2001)). Анализ таблицы показал, что в целом наилучшее соответствие данным дает прямое использование вероятностной модели, основанной на индикаторных свойствах видов.
Среди остальных моделей высокие результаты показала непараметрическая модель на основе топографического отображения, использующая ДДЗЗ. Этот результат получен даже несмотря на то, что в качестве факторов в ней не использовались характеристики рельефа, которые являются существенными для определения местообитаний видов. Обобщенные результаты для удобства представлены в таблице 5.1.
Таблица 5.1.
|
Оценка качества различных моделей |
||||
|
Вид |
Логистическая регрессия |
Нейронные сети прямого распространения |
Факторный анализ экологических ниш |
Модель на основе топографического отображения |
|
Белокрыльник болотный |
хорошее |
хорошее |
хорошее |
высокое |
|
Хамедафне прицветничковая |
хорошее |
хорошее |
хорошее |
высокое |
|
Ежа сборная |
хорошее |
хорошее |
хорошее |
Хорошее |
|
Кислица обыкновенная |
хорошее |
хорошее |
хорошее |
Хорошее |
|
Лютик кашубский |
хорошее |
хорошее |
хорошее |
Хорошее |
|
Ольха серая |
хорошее |
высокое |
хорошее |
высокое |
|
Пролесник многолетний |
хорошее |
высокое |
хорошее |
хорошее |
|
Прострел раскрытый |
хорошее |
хорошее |
хорошее |
высокое |
|
Страусник обыкновенный |
хорошее |
высокое |
хорошее |
хорошее |
|
Черника |
высокое |
высокое |
хорошее |
высокое |
Из таблицы видно, что модель на основе нейронных сетей прямого распространения и предложенная нами непараметрическая модель на основе топографического отображения позволяют добиться лучшего соответствия данным о присутствии видов по сравнению с другими моделями, в том числе и специально разработанными для моделирования местообитаний видов.
Для модели, использующей нейронные сети прямого распространения, это можно объяснить тем, что она позволяет моделировать более сложные нелинейные формы отклика видов на факторы среды.
Для непараметрической модели ее хорошее качество можно объяснить тем, что построенное топографическое отображение позволило неявно охарактеризовать свойства местообитаний без их содержательной интерпретации (что собственно и является преимуществом методов SOM и GTM) и извлечь из ДДЗЗ больше информации о сочетаниях факторов среды и о самих видах (распределение которых тоже отражено в спектральных характеристиках снимков), чем при традиционном способе оценки факторов среды по ДДЗЗ.
Таким образом, можно сделать вывод, что все рассмотренные нами модели показывают сравнимое качество и хорошие результаты для видов с различными экологическими характеристиками и широким диапазоном встречаемости на данной территории. Среди этих моделей предложенная нами модель на основе топографического отображения показывает стабильно более высокое качество и, кроме того, обладает рядом преимуществ, позволяющих рекомендовать ее для анализа и моделирования отклика на факторы среды как отдельных видов, так и единиц растительности.
Глава 6. Моделирование пространственного распределения видов флоры сосудистых растений
В данной главе обоснован комплексный подход к пространственному прогнозу распределения видов флоры, совмещающий учет отклика видов на градиенты факторов среды и моделирование стохастической составляющей геостатистическими методами.
Использование моделей отклика. Модели отклика отдельных видов флоры на факторы среды были использованы для вычисления для каждой ОТЕ регулярной сетки, охватывающей модельную территорию, вероятностной характеристики присутствия вида: для логистической модели по уравнению регрессии вычислялась вероятность присутствия вида; для нейросетевой модели вероятность присутствия вида вычислялась обученной нейронной сетью; для модели ФАЭН вычислялся индекс пригодности ОТЕ для вида; для непараметрическая модель на основе топографического отображения вероятность присутствия вида вычислялась по результатам калибровки. Полученные прогнозы были представлены в виде карт.
Прогноз логистической модели в большинстве случаев выглядит как сглаженный и “оптимистический” (с более высокими вероятностями) вариант прогноза нейросетевой модели, что объясняется общностью математического аппарата и большей гибкостью (числом степеней свободы) у нейронных сетей.
Сравнение модели ФАЭН и близкой к ней по задаче аппроксимации благоприятных для вида условий среды, но более гибкой модели на основе топографического отображения показывает, что для обильно представленных на территории видов результаты для обеих моделей весьма близки. Резкое различие результатов для мало распространенных видов (ольхи серой, белокрыльника болотного, хамедафне прицветничковой, ежи сборной) объясняется особенностями метода ФАЭН, который из-за недостатка информации не смог правильно оценить нишу вида, и сделав ее оценку слишком широкой, дал излишне “оптимистический” прогноз.
Сравнение пар моделей между собой показывает, что результаты (по крайней мере для наиболее распространенных видов), вполне сопоставимы, что согласуется с результатами сравнения моделей, приведенными в главе 5. Наиболее пространственно генерализованный результат получен для логистической модели, ограниченной монотонностью прогноза в пространстве факторов; наибольшую локальную пространственную вариабельность дает нейросетевая модель, имеющая максимальную гибкость; пространственная вариабельность моделей ФАЭН и топографического отображения примерно одинакова, и занимает промежуточное положение.
Таким образом, для целей картографического представления вероятностного распределения видов может быть рекомендована логистическая модель как наиболее генерализованная и отображающая общие закономерности их пространственного распределения. Модель ФАЭН нежелательно использовать для слабо распространенных на территории видов. Нейросетевая модель может быть рекомендована, если требуется наиболее детальная пространственная оценка распространения вида, однако ее применение ограничено опасностью адаптации нейронной сети к артефактам данных, т.е. требованием использования большого объема данных. Модель на основе топографического отображения представляется сбалансированной по степени генерализации и требованиям к объему исходных данных, особенно если недоступны полевые данные о факторах среды (эта модель использует только данные дистанционного зондирования Земли).
Анализ стохастической пространственной составляющей. В качестве модели тренда для вероятности присутствия выбранных видов были использованы прогнозы логистической модели. Остатки от тренда вычислялись вычитанием его из реальной вероятности присутствия вида, которая оценивалась по соседним площадкам как относительная встречаемость вида в небольшой окрестности данной площадки описания.
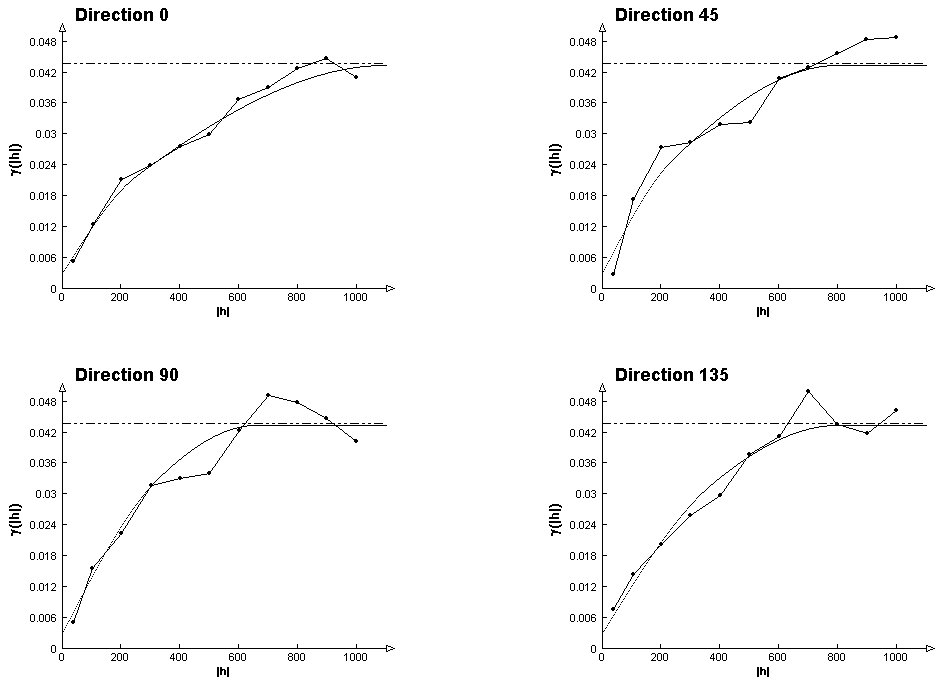
Для всей видов было установлено соответствие полученных остатков требованиям, предъявляемым к реализации случайных функций, проведен вариограммный анализ, построены модели пространственных зависимостей (вариограммы), и выполнено моделирование значений остатков для всей ОТЕ методом кригинга. Детальное математическое описание использованных геостатистических методов можно найти в (Cresse, 1993). Для вариограммного анализа использовалась программа VarioWin v2.1. На рис.6.1 даны графики теоретических и экспериментальных вариограмм остатков по направлениям для черники, на которых видно их хорошее согласование.
|
|
|
Рис.6.1. Графики экспериментальной и теоретической вариограмм остатков по направлениям для черники. |
Для выбранных видов были построены модели вариограмм остатков, наилучшим образом приближенные к экспериментальным вариограммам – теоретические вариограммы. Параметры моделей подбирались в интерактивном режиме.
В таблице 6.1 дано описание соответствующих моделей вариограмм остатков и полученные значения параметров этих моделей.
Таблица 6.1.
Модели вариограмм остатков
|
Вид (код вида) |
Эффект самородков |
Тип базисных функций |
Ранг (м) |
Порог |
Угол (градус) |
Анизотропия |
|
Черника (945) |
0.003 |
Сферическая |
390 |
0.00800 |
120 |
0.6 |
|
Сферическая |
1000 |
0.03250 |
0 |
0.6 |
||
|
Кислица обыкновенная (333) |
Сферическая |
640 |
0.05000 |
0 |
0.7 |
|
|
Страусник обыкновенный (857) |
Сферическая |
580 |
0.00900 |
0 |
0.8 |
|
|
Сферическая |
1000 |
0.02380 |
105 |
0.7 |
||
|
Пролесник многолетний (730) |
Сферическая |
500 |
0.00600 |
0 |
1 |
|
|
Сферическая |
1000 |
0.04400 |
30 |
0.6 |
||
|
Лютик кашубский (468) |
0.0006 |
Сферическая |
700 |
0.02580 |
350 |
0.6 |
|
Прострел раскрытый (739) |
0.0008 |
Сферическая |
440 |
0.00900 |
0 |
0.6 |
|
Сферическая |
1000 |
0.01240 |
90 |
0.7 |
||
|
Ольха серая (587) |
0.0003 |
Сферическая |
660 |
0.00090 |
135 |
0.6 |
|
Сферическая |
730 |
0.00600 |
0 |
1 |
||
|
Белокрыльник болотный (24) |
Сферическая |
400 |
0.00100 |
100 |
0.6 |
|
|
Сферическая |
500 |
0.00510 |
0 |
0.7 |
||
|
Хамедафне прицветничковая (40) |
Сферическая |
250 |
0.00095 |
0 |
1 |
|
|
Сферическая |
400 |
0.00330 |
0 |
2.5 |
||
|
Ежа сборная (228) |
Сферическая |
390 |
0.01520 |
0 |
1.2 |
|
Моделирование стохастической составляющей. Для выбранных видов и для всех ОТЕ на модельной территории с использованием полученных вариограмм было выполнено моделирование методом кригинга остатков (отклонений от тренда, заданного логистической регрессией). Оценка результатов методом перекрестной проверки показала, что полученные пространственные оценки остатков объясняют от 65 до 85% изменчивости данных (стохастической составляющей), что является свидетельством хорошего качества результатов. Для вычислений использовалась программа Surfer v8.0.
Кроме этого, была выполнена симуляция остатков на модельную территорию, позволяющая воспроизвести не только средние значения, но и характеристики пространственной изменчивости стохастической составляющей. Симуляция осуществлялась двумя методами, с использованием ранее полученных моделей вариограмм – для Гауссовой симуляции использовалась программа SGSIM из пакета программ GSLib (Deutsch, 1998); для искусственного отжига – программа SASIM из того же пакета.
Как и следовало ожидать, метод кригинга оказался наиболее “консервативным”, он не корректирует логистическую модель в тех областях пространства, где недостаточно данных, имеет наименьшие отклонения от фактических значений, и дает наиболее сглаженную пространственную картину. Для основанного на нем метода Гауссовой симуляции оценки остатков могут отклоняться от полученных кригингом в любую сторону при сохранении средних значений. Кроме того, данный метод воспроизводит стохастическую пространственную составляющую (точнее, характер ее пространственного распределения, а не точные пространственные значения) и там, где данных для кригинга недостаточно. Метод искусственного отжига еще больше ориентирован на воспроизведение характеристик (статистик), а не значений, поэтому он может быть рекомендован скорее для генерации искусственных данных, распределение которых в пространстве совпадает с требуемым, а не для коррекции моделей реального распределения видов.
Полученные модели остатков складывались с трендом (прогнозом логистической модели) для получения прогноза пространственного распределения видов на всей территории, учитывающего не только факторы среды, но и данные о присутствии видов из БД “Флора”.
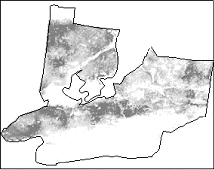
Наконец, полученные прогнозы были использованы для картографической оценки ареалов распространения видов (пространственной классификации) на основе теории Марковских случайных полей методом ICM (Besag, 1984).
|
|
|
|
Вероятность присутствия |
Потенциальный ареал |
|
Рис.6.2. Построение потенциального ареала распространения черники |
|
Метод позволяет не только учесть вероятности присутствия вида в ОТЕ, но ее и пространственный контекст (присутствие вида в соседних ОТЕ), а также получить ареал заданной пространственной генерализации. На рис.6.2. для иллюстрации приведены вероятность присутствия черники на модельной территории, полученная с учетом стохастической составляющей, и построенный на ее основе ареал.
Выводы
.
Список основных публикаций
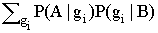 .
.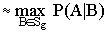 .
.