| Выпуск 1
|
Выпуск 3
|
Конец |
Список
|
ПРАКТИКУМ ПО МАТЕМАТИЧЕСКИМ МЕТОДАМ В ЭКОЛОГИИ:
Выпуск 2
С.С. Крамаренко
МЕТОДЫ ОЦЕНКИ АБСОЛЮТНОЙ ЧИСЛЕННОСТИ
Введение
1. Методы оценки абсолютной численности, основанные на использовании индексов плотности
Введение
Во второй части практикума рассматриваются вопросы оценки абсолютной численности особей популяции. Большая часть этих методик основана на подходе, который называется “мечение – выпуск – повторный отлов” (МВПО) (capture-recapture model). В его основе лежит предположении, что меченое животное имеет такую же вероятность быть отловленным, как и немеченое.
В общем случае мечение представляет собой процедуру присвоения организму определенного признака (метки), отличающего его от другого (немеченого) животного. Метки могут быть либо индивидуальными (кодово-номерными), либо унифицированными (единообразными) для группы животных в целом.
Важными особенностями как самой метки, так и всей процедуры мечения в целом, являются следующие:
— процедура мечения должна быть относительно проста и не оказывать отрицательного влияния на все процессы жизнедеятельности меченого организма;
— метки должны быть относительно долговечными (идеальный вариант, когда метка сохраняется на протяжении всей жизни животного);
— сами метки не должны демаскировать животное и повышать вероятность быть обнаруженным хищниками;
— наличие метки не должно оказывать влияние на вероятность отлова данной особи.
Примеры меток широко известны — это кольцевание птиц, летучих мышей, рыб, отрезание пальцев у грызунов, радиометки у крупных млекопитающих, нанесение пятен несмывающейся краски у некоторых беспозвоночных и т.п.
Методы МВПО используются чаще всего для относительно крупных организмов с продолжительным жизненным циклом, чью численность или плотность невозможно оценить другими способами. Помимо оценки численности данная группа методов также может быть использована для оценки целого ряда экологических характеристик популяции, а именно:
1. Уровня подвижности организмов и определения границ индивидуального участка.
2. Скорости индивидуального (хронологического) роста.
3. Демографических показателей и т.п.
Методы анализа миграционной активности и территориальности особей, роста организмов (хронологического и аллометрического), демографии, динамики численности популяций и т.п. будут рассмотрены нами в следующих выпусках практикума.
1. Методы оценки абсолютной численности, основанные на использовании индексов плотности
Индексом плотности называется любая однозначно и четко регистрируемая величина, эквивалентно связанная с численностью особей. Индексом плотности может быть и число фекалий животных на единицу маршрута, и количество силуэтов птиц, появляющихся на фоне диска Луны за единицу времени, и любая другая оценка плотности популяции линейно связанная с ее численностью.
Именно обязательное соблюдение наличия прямолинейной связи между индексом плотности и абсолютной численностью популяции лежит в основе двух предложенных ниже методов оценки ее численности. Потому что в этом случае можно ожидать, что при изъятии или добавлении особей в исследуемой популяции индекс плотности будет пропорционально снижаться или возрастать, соответственно.
Для получения достоверных результатов необходимо соблюдать ряд важных требований:
1. Границы участка обязательно должны быть четкие и непроницаемые для животных.
2. Смертность и рождаемость между периодами исследования должны быть по возможности минимальными.
3. Практически должна отсутствовать эмиграции и иммиграция животных.
4. Промежуток времени между обследованиями должен быть по возможности минимальным.
5. Достаточно высокой должна быть точность оценок индекса плотности.
1.1. Метод двукратного использования индексов
Данный метод заключается в следующем. В конкретных условиях определяется индекс плотности (
i1) популяции. Затем к этой популяции добавляется (или из нее изымается) известное количество животных (C). Определяется новое значение индекса плотности (i2).Оценка абсолютной численности животных тогда может быть определена по формуле (Коли, 1979):
|
|
(1.1) |
Если животные добавляются, то величина
C входит в формулу 1.1 со знаком плюс (+), если изымаются — со знаком минус (-).1.2. Метод Келкера
Если популяция сама оказывается достаточно гетерогенной (т.е. в ней можно выделить два или более четко различимых класса по окраске, по полу, по возрасту и т.п.), то численность такой популяции может быть определена, используя метод селективного изъятия (или добавления) Келкера (Kelker, 1940; 1944; Коли, 1979).
Данный метод может быть использован не только при наличии двух (или более) классов среди животных исследуемой популяции, но также и при наличии двух (или более) видов, совместно обитающих на одной территории (они должны быть экологически сходными).
Последовательность этапов определения абсолютной численности с использованием метода Келкера заключается в следующем.
1. Вначале бескровным образом отлавливается выборка животных и в ней определяется относительная доля особей каждого класса.
2. Далее из этой популяции изымается (или к ней добавляется) фиксированное количество особей одного из классов (или обоих классов).
3. Отлавливается новая выборка и в ней снова определяется относительная доля особей каждого класса.
4. Исходную численность популяции можно тогда определить по формуле:
|
|
(1.2) |
где
X, Y — два класса животных в популяции; N1 — численность популяции при первом учете; p1 — доля особей, принадлежащих классу X при первом учете; p2 — доля особей, принадлежащих классу X при втором учете; CX, CY — число особей классов X и Y, соответственно, добавляемое (или изымаемое) из популяции между двумя последовательными обследованиями. Если особи добавляются, то CX, CY — положительные величины, если изымаются — отрицательные. Если особи какого-то из классов не добавляются и не изымаются, то эта величина равна нулю. Величина C = CX - CY.Данный метод нельзя использовать в тех случаях, если особи в популяции не распределены равномерно.
2. Метод неселективного изъятия
Данный метод используется для оценки абсолютной численности животных, обитающих на ограниченной территории. Он может быть применен для оценки численности насекомых на определенном участке луга, млекопитающих в локальной популяции и т.п. В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности встречаемости животного в серии последовательных отловов, вызванное снижением численности популяции в результате изъятия из нее особей.
Таким образом, при применении данного метода животных отлавливают, подсчитывают их количество и не выпускают до конца исследования. Затем производят еще 3-4 последовательных отлова по аналогичной методике, при этом число отловленных животных постепенно уменьшается, вследствие уменьшения их общего количества на исследуемой территории. Если теперь построить график зависимости числа отловленных животных при каждом отлове от общего числа ранее отловленных, то по нему можно найти оценку исходной численности популяции (Грин и др., 1990).
Пример
. В серии четырех последовательных отловов было зарегистрировано 120, 93, 60 и 35 животных, соответственно. Необходимо оценить исходный уровень численности популяции.Таблица 1
|
Номер |
Количество отловленных |
Совокупное количество ранее |
|
1 |
120 |
0 |
|
2 |
93 |
0 + 120 = 120 |
|
3 |
60 |
0 + 120 + 93 = 213 |
|
4 |
35 |
0 + 120 + 93 + 60 = 273 |
Используя любой статистический пакет (например, STATISTICA), находим параметры уравнения линейной зависимости: Y = a – b·X:
a = 123,9 + 5,5,
b = 0,31 + 0,03.
Искомая величина находится из условия, что
Y = 0.N = 123,9/0,31 = 399,6 @ 400 особей.
На рисунке 1 это значение соответствует точке пересечения линии регрессии с осью OX.
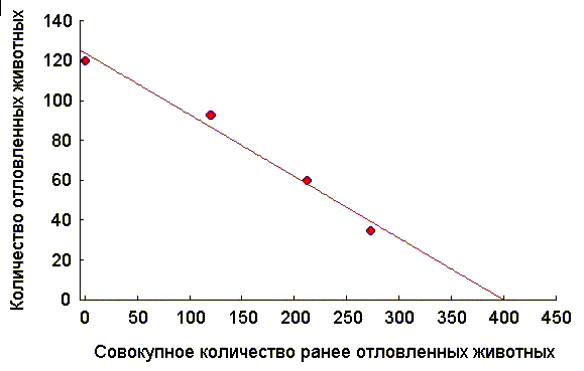
Рис. 1
Коэффициент
b данного уравнения дает оценку средней улавливаемости особей, т.е. доли отловленных особей от их исходной численности.Например, если изначально было 400 особей (как мы рассчитали), то во время первого отлова теоретически должно быть отловлено:
N1
= N0 ·b = 400·0,31 = 124 особи,во время второго отлова:
N2
= (N0 - N1)·b = (400 - 124)·0,31 » 86 особей,во время третьего:
N3
= (N0 - N1 - N2)·b = (400 - 124 - 86)·0,31 » 59 особей,и т.д.
Поскольку во время первого отлова мы отловили часть особей, то естественно исходная численность популяции перед проведением второго отлова снизилась на эту величину.
Однако довольно часто встречаются случаи, когда особи обладают высокой миграционной активностью и место отловленных резидентных особей сразу же занимают иммигранты. (Подобная ситуация может встретиться, например, при отловах мелких млекопитающих.) Тогда совокупное число отловленных животных не будет снижаться при каждом последующем отлове, что даст заниженную оценку коэффициента
b и, следовательно, завысит оценку начальной численности популяции.В таких случаях уравнение зависимости совокупного количества ранее отловленных животных от количества отловленных во время каждого последовательного отлова модифицируется и в него включается новый показатель, характеризующий интенсивность миграции (Лукьянов, 1988):
|
Y = a - b·X + m·(n - 1), |
(2.1) |
где n — номер отлова. Коэффициент
b уравнения 2.1 также даст оценку средней улавливаемости.Тогда исходную численность резидентных особей можно рассчитать по формуле:
|
|
(2.2) |
а оценку потока иммигрантов:
|
|
(2.3) |
Совокупная численность оседлых и мигрантов тогда будет равна:
|
I = N0 + M, |
(2.4) |
а доля мигрантов в ней:
|
|
(2.5) |
3. Методы мечения, выпуска и повторного отлова (МВПО)
3.1. Метод Петерсена-Линкольна
Наиболее известным и широко распространенным из данной группы методов является метод Петерсена-Линкольна, который был модифицирован Бейли (
Petersen, 1896; Lincoln, 1930; Bailey, 1951; 1952; Poole, 1974; Begon, 1979; Коли, 1979). Он заключается в следующем. Отлавливается выборка животных численностью M, каждое животное метится и выпускается обратно. Спустя некоторое время отлавливается новая выборка животных объемом n и среди них отмечается количество ранее меченых (m). Тогда исходную численность популяции можно оценить по формуле:|
|
(3.1) |
Ошибка оценки численности составляет:
|
|
(3.2) |
Важным условием использования данного метода является продолжительность интервала между двумя последовательными отловами - он должен быть минимальным для того, чтобы избежать влияния процессов рождения и миграции на получаемый результат. С другой стороны, он должен дать возможность меченым особям более или менее равномерно рассредоточиться среди остальных особей популяции.
Смертность особей между двумя момента отлова не смещают оценки исходной численности популяции
N.3.2. Метод Бейли
Метод трехкратных отловов Бейли
позволяет, помимо оценки численности популяции, оценить также показатели рождаемости и смертности в течение периода исследования (Bailey, 1951; 1952; Коли, 1979). Кроме того, он применим в том случае, если животные довольно активно перемещаются.Для того чтобы оценить численность популяции с помощью метода Бейли необходимы иметь данные по двум последовательным мечениям и двум последовательным отловам. Таким образом, в момент
t0 производится отлов выборки животных. Их всех единообразно метят и выпускают обратно. Спустя некоторое время в момент t1 отлавливается новая выборка, в ней подсчитывается количество меченых особей. Остальные (немеченые) животные из этой выборки также метятся, но на этот раз новой меткой, отличной от первой. Спустя время в момент t2 вновь отлавливается выборка животных и в ней подсчитывается число особей, меченых в первый раз и во второй.Оценка численности популяции и ее ошибки в момент
t1 тогда составляет:|
|
(3.3) |
|
|
(3.4) |
В промежутке между моментами
t1 и t2 скорость увеличения численности популяции за счет появления новорожденных особей и иммиграции определяется по формуле:|
|
(3.5) |
а скорость убыли популяции в промежутке между моментами
t0 и t1 за счет гибели особей и эмиграции:|
|
(3.6) |
Так как продолжительность временных интервалов между моментами
t0 и t1, с одной стороны, и t1 и t2, с другой, часто бывают различными, прежде чем сравнивать эти скорости, необходимо привести их к эквивалентной продолжительности времени:|
|
(3.7) |
|
|
(3.8) |
Полагая, что эти скорости остаются постоянными на протяжении всего периода исследования, можно определить скорость роста численности популяции за этот промежуток времени:
r = b - d.Тогда численность популяции в момент времени
t0 составляет:|
|
(3.9) |
а в момент времени
t2:|
|
(3.10) |
Пример.
Был произведен трехкратный отлов животных с последовательными мечениями (табл. 2).Необходимо определить численность популяции в каждый момент времени, если второй отлов производили через 4 недели после первого, а третий — через 6 недель после второго.
Таблица 2
|
Время (t) |
Количество меченых животных |
Количество животных отловленных для повторного мечения |
Отловлено из числа M0 |
Отловлено из числа M1 |
|
0 |
M 0 = 600 |
|||
|
1 |
M 1 = 518 |
n 1 = 700 |
m 01 = 182 |
|
|
2 |
n 2 = 700 |
m 02 = 146 |
m 12 = 158 |
Вначале, используя формулы 3.3 и 3.4, определим оценку численности популяции в момент времени
t1 и ее ошибку:N1
=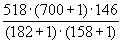 = 1822 особей,
= 1822 особей,
SEN1 = 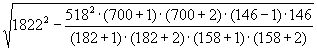 = 238
= 238
Затем определим величины изменения численности популяции:
B12
=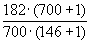 = 1,240
= 1,240
и
D01
=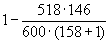 = 1 - 0,793 = 0,207.
= 1 - 0,793 = 0,207.
Поскольку первый промежуток времени составляет 4 недели, а второй — 6, то приводим все показатели к однонедельному отрезку времени:
b12
= 0,215 : 4 = 0,0358,d01 = 0,233 : 6 = 0,0583.
Скорость роста численности данной популяции за анализируемый отрезок времени тогда составляет:
r = 0,0358 - 0,0583 = -0,0225.Наконец, определяем численность популяции в момент времени
t0: N0 =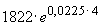 = 1994 особей и t2: N2 =
= 1994 особей и t2: N2 = 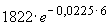 = 1592 особей.
= 1592 особей.
3.3. Метод многократного отлова и мечения
Оценить численность популяции можно и проводя многократные отловы, при этом производя единообразное мечения всех ранее немеченых животных при каждом последовательном отлове (
Poole, 1974). В этом случае можно ожидать, что с каждым последовательным отловом и мечением в популяции будет расти доля меченых особей вплоть до того момента, когда все особи в данной популяции окажутся с метками. Таким образом, на основании оценки доли меченых особей в популяции (полученной с помощью выборочных данных) в зависимости от совокупного числа всех ранее меченых животных можно найти оценку величины популяции – ее численность, при которой доля меченых животных должна достичь 100 %.Пример.
Было произведено шести последовательных отловов, при этом, каждый раз производилось мечение (меткой одного и того же типа) всех ранее немеченых животных.Все полученные результаты и промежуточные расчеты занесены в таблицу 3.
Таблица 3
|
Номер отловов ( i) |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
||
|
Кол-во отловленных ( ni) |
58 |
58 |
42 |
41 |
33 |
38 |
|
|
Кол-во вновь меченых из числа отловленных ( mi) |
— |
42 |
18 |
16 |
13 |
8 |
|
|
Число ранее меченых |
— |
16 |
24 |
25 |
20 |
30 |
|
|
Совокупное число ранее меченых ( Mi) |
— |
58 |
100 |
118 |
134 |
147 |
|
|
Доля ранее меченых среди отловленных ( pi) |
— |
27,6 |
57,1 |
61,0 |
60,6 |
78,9 |
|
Таким образом, в первый раз отловили 58 животных, всех их пометили и отпустили. Во второй раз также отловили 58 животных, из них 16 имели метки (т.е. 16·100/58 = 27,6 %), а 42 пометили впервые. Совокупное число ранее меченых особей, таким образом, составляет 58 особей. В третий раз отловили 42 животных. Меченых из них было 24 (т.е. 24·100/42 = 57,1 %), а 18 пометили впервые. Теперь совокупное число меченых особей составляет 58+42 = 100 особей. И так далее.
Если теперь построить уравнение зависимости доли меченых организмов от совокупного числа меченых (рис. 2), то полученная зависимость может быть описана уравнением регрессии:
Y
= 0,1 + 0,51·X.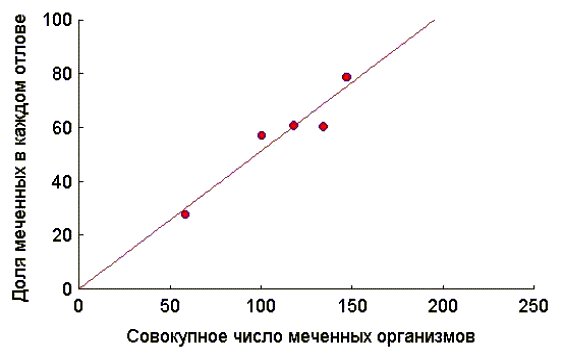
Рис. 2
Нас интересует значение
X, при котором Y = 100, т.е. при какой численности все особи в выборке окажутся уже ранее мечеными.Также можно проверить справедливость предположения о постоянстве вероятности отлова для всех особей. В этом случае линия регрессии (рис. 2) должна проходить через начало координат и иметь наклон 100/
N, если мы используем процентную форму записи доли меченых особей в каждом отлове (или 1/N, если частота меченных животных отмечается в долях единицы).Можно также оценить и среднюю долю особей, учитываемых при каждом последовательном отлове. Для этого среднюю за несколько последовательных отловов численность животных необходимо поделить на полученную оценку численности популяции. Среднее число отлавливаемых животных в нашем примере составляет 270 : 6 = 45. Отношение этой величины к оценке популяции составляет: 45 : 229 = 0,197. Таким образом, в среднем отлавливается только каждое пятое животное. И
если на другом сходном участке или в другой момент времени мы отловим 65 особей, то можно сделать вывод, что численность этой популяции составляет около 330 особей.3.4. Метод Шумахера и взвешенной средней
На принципе, изложенном выше основаны также и два других метода — метод Шумахера и метод взвешенной средней
.Оценка численности популяции согласно методу Шумахера составляет (
Schumacher, Eschmeyer, 1943):|
|
(3.11) |
а согласно методу взвешенной средней (
Begon, 1979):|
|
(3.12) |
где обозначения те же, что и в таблице 3.
Пример. Рассчитаем оценки численности популяции, используя метод Шумахера и метод взвешенной средней для данных, приведенных в таблице 3.
Для удобства расчетов составим новую таблицу 4.
Таблица 4
|
Показатели |
Номера отловов ( i) |
Суммы |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
|||
|
ni |
58 |
58 |
42 |
41 |
33 |
38 |
270 |
|
|
mi |
— |
42 |
18 |
16 |
13 |
8 |
97 |
|
|
Mi |
— |
58 |
100 |
118 |
134 |
147 |
— |
|
|
Mi*ni |
— |
3364 |
4200 |
4838 |
4422 |
5586 |
22410 |
|
|
Mi2*ni |
— |
195112 |
420000 |
570884 |
592548 |
821142 |
2599686 |
|
|
Mi*mi |
— |
2436 |
1800 |
1888 |
1742 |
1176 |
9042 |
|
Используя промежуточные результаты расчетов из таблицы 4, находим искомые оценки:
NSh
= 2599686 : 9042 = 288 особей,NWM
= 22410 : (97+1) = 229 особей.Реальное значение численности данной популяции, оцененное на основе 12 последовательных отловов составляет 243 особи. Таким образом, можно отметить, что метод Шумахера дает несколько завышенные оценки численности популяции, по сравнению с методом взвешенной средней.
3.5. Метод Джолли-Зебера
Если при каждом последовательном отлове особям присваиваются индивидуальные метки, то в этом случае можно оценить численность популяции (а также целый набор других экологических популяционных показателей) на основе метода Джолли-Зебера
(Jolly, 1965; 1982; Seber, 1965; Begon, 1979; Коли, 1979).Использование этого метода требует, чтобы в распоряжении исследователя были подробные сведения относительно дат мечения и отловов каждого конкретного животного. Обычно для данных, полученных при
i-том отлове, используют следующие обозначения:|
Ni |
— оценка численности популяции; |
|
ni |
— размер выборки; |
|
Mi |
— общее число животных, меченых в популяции до i -того отлова; |
|
mi |
— число меченых животных в выборке; |
|
Ri |
— число меченых и отпущенных на волю животных; |
|
ri |
— число животных из числа Ri, выпущенных на волю после i-того отлова и затем снова попавшихся при последующих отловах; |
|
Zi |
— число животных, помеченных до i-того отлова, не попавшихся при i-том отлове, но попавшихся при одном из последующих; |
|
ai |
— доля меченых животных среди попавшихся i-том отлове. |
Особенности использования данного примера рассмотрим на примере, приведенном в книге Г.Коли
(1979).Результаты шести последовательных отловов занесены в таблицу 5.
Таблица 5
|
i |
ni |
Ri |
Число раз, которое животные попадались прежде |
mi |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
||||
|
1 |
9 |
9 |
— |
— |
|||||
|
2 |
13 |
12 |
5 |
5 |
|||||
|
3 |
14 |
14 |
2 |
7 |
9 |
||||
|
4 |
8 |
7 |
0 |
2 |
1 |
3 |
|||
|
5 |
18 |
16 |
1 |
2 |
3 |
1 |
7 |
||
|
6 |
14 |
14 |
0 |
1 |
2 |
1 |
2 |
— |
6 |
|
ri |
8 |
12 |
6 |
2 |
2 |
— |
|||
Для любого отлова 1 <
i < k оценка численности популяции рассчитывается по формуле:|
|
(3.13) |
а ее ошибка:
|
|
(3.14) |
где
|
|
(3.15) |
|
|
(3.16) |
Составим вспомогательную
таблицу 6, которая помогает понять расчет показателей Zi.Таблица 6
|
m2 |
5 |
||||
|
m3 |
2 |
2+7=9 |
|||
|
m4 |
0 |
0+2=2 |
0+2+1=3 |
||
|
m5 |
1 |
1+2=3 |
1+2+3=6 |
1+2+3+1=7 |
|
|
m6 |
0 |
0+1=1 |
0+1+2=3 |
0+1+2+1=4 |
0+1+2+1+2=6 |
|
2+0+1+0=3 |
2+3+1=6 |
6+3=9 |
4 |
— |
|
|
Z2 |
Z3 |
Z4 |
Z5 |
Z6 |
Тогда, например, в момент времени
i = 2 оценка численности данной популяции составит: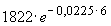 особей.
особей.
Ошибка этой оценки в свою очередь будет равна:
|
|
|
|
Оценка
N1 рассчитываются на основе показателя скорости роста численности популяции:|
r = |
(3.17) |
|
|
(3.18) |
Оценка численности популяции во время проведения последнего отлова
Nk , в свою очередь, также может быть получена с использованием соответствующего показателя скорости роста численности популяции (3.17).В тех случаях, когда постановка эксперимента предусматривает лишь троекратное проведение отловов, метод Джолли-Зебера может быть использован наряду с методом трехкратных отловов Бейли.
Литература
Грин Н., Стаут У., Тейлор Д
. Биология: В 3-х т. Пер. с англ. – М.: Мир, 1990.Коли Г. Анализ популяций позвоночных. Пер. с англ.
– М.: Мир, 1979. – 364 с.Лукьянов О.А. Оценка демографических параметров популяций мелких млекопитающих методом безвозвратного изъятия // Экология.
– 1988. – № 1. – С.47-55.Bailey N
.T.J. On estimating the size of mobile population from recapture data // Biometrika. – 1951. – V.38. – P.293-306.Bailey N.T.J. Improvements in the interpretation of recapture data // J.Anim.Ecol.– 1952. – V.21. – P.120-127.
Begon M. Investigating animal abundance: Capture-recapture for biologists. – Baltimore: University Park Press, 1979. – 97 p.
Jolly G.M. Explicit estimates from capture-recapture data with both death and immigration – stochastic model // Biometrika. – 1965. – V.52. – P.225-247.
Jolly G.M. Mark-recapture models with parameters constant in time // Biometrics. – 1982. – V.38. – P.301-321.
Kelker G.H. Estimating deer populations by differential hunting loss in the sexes // Proc.Utah.Acad.Sci.Arts and Letters. – 1940. – V.17. – P.65-69.
Kelker G.H. Sex ratio equations and formulas for determing wildlife populations // Proc.Utah.Acad.Sci.Arts and Letters. – 1944. – V.20. – P.189-198.
Lincoln F.C. Calculating waterfowl abundance on the basis of banding returns // Cir.U.S.Dept.Agric. – 1930. – No. 118.
Petersen C.G.J. The yearly immigration of young plaice into Limfjord from the German sea etc. // Rept.Danish Biol.Stn. – 1896. – V.6. – P.1-48.
Poole R.W. An introduction to quantitative ecology. – New-York: McGraw-Hill, 1974. – 532 p.
Schumacher F.X., Eschmeyer R.W. The estimation of fish populations in lakes and ponds // J.Tenn.Acad.Sci. – 1943. – V.18. – P.228-249.
Seber G.A.F. A note on thy multiple-recapture census // Biometrika. – 1965. – V.52. – P.249-259.
| ‚ыпуск 1
|
Выпуск 3
|
Начало |
Список
|
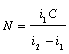
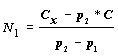 ,
,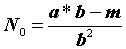 ,
,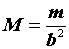 .
.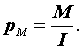
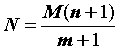 .
.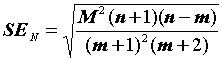 .
.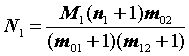 ,
,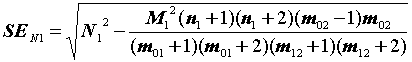
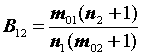 ,
,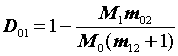 .
.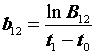 ,
,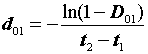 .
.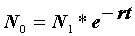 ,
,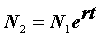 .
.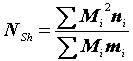 ,
,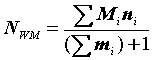 ,
,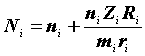 ,
,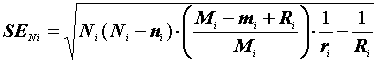 ,
,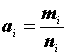 ,
,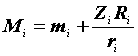 .
.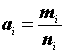 ,
,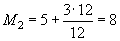 ,
,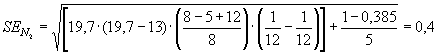 .
.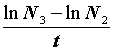 ,
,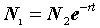 .
.