| Выпуск 2 |
Конец |
Список
|
ПРАКТИКУМ ПО МАТЕМАТИЧЕСКИМ МЕТОДАМ В ЭКОЛОГИИ:
Выпуск 3
С.С. Крамаренко
МЕТОДЫ АНАЛИЗА ВОЗРАСТНОЙ СТРУКТУРЫ ПОПУЛЯЦИИ
Введение
1. Метод выделения когорт Хазена-Хардинга
2. Оценка возрастного разнообразия
3. Таблицы выживания и особенности их построения
Литература
Введение
Любой организм рождается, живет и в конце своего жизненного пути умирает. За отведенный ему срок он сначала растет, затем по достижению половой зрелости вступает в размножение (если, конечно, повезет найти полового партнера), стараясь оставить после себя как можно больше потомков. Процессы роста, дифференциации и репродукции, естественно, требуют определенных энергетических и вещественных затрат, причем на разных стадиях жизненного цикла животным необходимо разное количество энергии и минеральных веществ. С другой стороны различные возрастные группы потребляют пищу разного качества и в различном количестве, занимая при этом различные места в цепи питания (например, личинки бабочек питаются грубой растительной пищей, а имаго — нектаром цветов; молодь грызунов выкармливается молоком матери, а адультные особи — растительноядны; и т.п.). Следовательно, особи разного возраста различаются в отношении потока энергии, проходящей через популяцию.
На разных стадиях онтогенеза особи имеют и различные миграционные характеристики (молодь более седентарна, чем достигшие половой зрелости и находящиеся в поиске брачного партнера молодые самцы) и, таким образом, также различаются по степени и направленности переноса вещества, энергии и информации (генетической) в популяции и биогеоценозе в целом. Соотношение различных возрастных групп также в значительной мере определяет репродуктивную стратегию вида, динамику численности (через соотношение рождаемости и смертности особей различных возрастных групп; см. ниже), пространственную и фенотипическую структуры популяции и т.д. Следовательно, возрастная структура популяции, т.е. соотношение особей различного возраста в популяции, является немаловажной характеристикой и разносторонне должна быть изучена при проведение исследований по популяционной экологии.
Дифференциация особей по возрастам может быть произведена, используя или геохронологические единицы (сутки или год, и их производные — неделю, декаду, месяц, квартал, сезон года), или биохронологические (стадии жизненного цикла, продолжительность которых может быть различной в геохронологических единицах). Первый подход используется, как правило, для организмов с детерминированным ростом, т.е. когда взрослая особь — это аллометрически увеличенная копия молодой, а второй — для животных, претерпевающих метаморфозы в онтогенезе (например, у насекомых с полным превращением можно выделить четыре основные возрастные стадии — яйцо, личинку, куколку и имаго). При этом различия в общей продолжительности жизни определяют период генерации, который в одних случаях может быть меньше стандартной геохронологической единицы — года (например, у мелких млекопитающих и многих насекомых за год сменяется три или четыре поколения), или намного превышать ее (у крупных млекопитающих средняя продолжительность генерации составляет десятки лет). Наиболее простейшим вариантом возрастной дифференциации может быть выделение в популяции особей предрепродукционной, репродукционной и пострепродукционной стадий (Яблоков, 1987). В этом случае, основным показателем отнесение особи к тому или иному возрастному классу является ее способность к воспроизведению.
При использовании геохронологических единиц основная задача исследователя заключается в определении с заданной степенью точности соответствия между размерами организма (или его весом) и возрастом, измеренным в сутках (месяцах, годах или др.). Это можно сделать проследив за хронологическим ростом группы новорожденных особей, появившихся в течение фиксированного момента времени (когортой). Правда этот метод плохо “срабатывает” при определении возраста половозрелых особей, рост тела которых практически завершен. Тогда используют видоспецифические особенности, позволяющие определить возраст таких животных (например, по степени стертости зубов у многих грызунов, или по количеству слоев эмали на зубах у ластоногих, или по количеству линий нарастания на чешуе рыб, и т.п.).
Часто просто разбивают популяцию на возрастные группы, используя лишь размеры (или вес) особей. В таком случае ведут речь о размерно-возрастном распределении, и используется оно, как правило, для организмов с детерминированным ростом. Понятие “возраста”, таким образом, заменяется понятием “размер”.
1. Метод выделения когорт Хазена-Хардинга
Возрастное распределение представляет собой одномоментный “срез” через популяцию. На рисунке 1 схематически изображены кривые роста размеров тела особей из разных когорт (продолжительность генерации составляет 2 единицы времени).
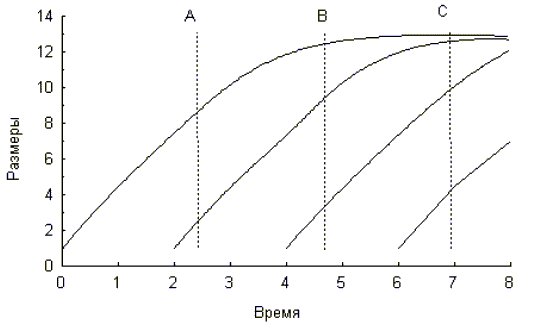
Рис. 1
В зависимости от времени проведения анализа размерно-возрастного распределения популяция может быть более или менее гетерогенна в отношение возраста. Например, в момент времени A, в популяции можно отметить присутствие особей двух возрастных (и размерных) групп, в момент времени B — трех, а в момент времени C — всех четырех, хотя как говорилось выше, очень трудно провести дифференциацию двух старших возрастных групп, поскольку их размеры очень близки (рис.1).
Графическим представлением размерно-возрастной структуры популяции являются частотные гистограммы распределения. На рисунке 2 представлена гистограмма размерно-возрастного распределения моллюска Helix albescens из одной локальной популяции г. Симферополя (Крым, Украина) на основании размеров большого диаметра раковины. Явно просматривается наличие трех когорт.
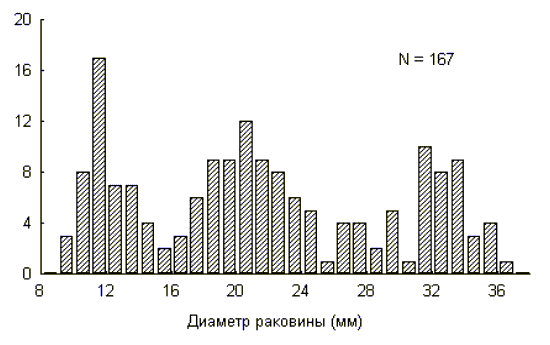
Рис. 2
Популяция, таким образом, представляет собой смесь различных когорт. Так как распределение особей в каждой когорте, как правило, имеет нормальное распределение (см. рис. 2), то можно определить число когорт, а также принадлежность каждой особи к той или иной когорте, используя нормальную вероятностную бумагу (Hazen, 1913; Harding, 1949).
Как известно, при изображении значений выборки, имеющей нормальное распределение, на нормальной вероятностной бумаге, через точки можно провести прямую линию. И эта особенность нормальной вероятностной бумаги часто используется при проверке соответствия данного распределения нормальному (Компьютерная …, 1990; Справочник …, 1989). В этом случае особи, принадлежащие разным когортам будут располагаться на прямых, имеющих различный наклон. Для данных, по наземному моллюску Helix albescens распределение точек на нормальной вероятностной бумаге приведено на рис. 3.
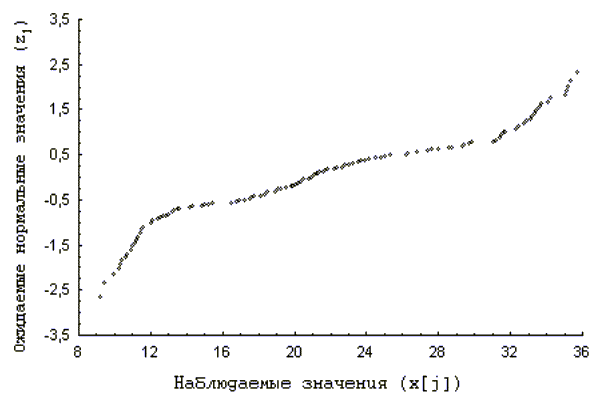
Рис. 3
Построение таких скаттеров вручную при большом объеме выборки робота довольно хлопотная, поэтому можно рекомендовать использовать соответствующую опцию (Normal probability plot), которая имеется во многих пакетах статистических прикладных программ, например STATISTICA. Алгоритм ее следующий.
Вначале все значения выборки ранжируются от самой меньшей до самой большей; теперь каждое значение представляется в виде x[j], где j – это ее порядковый номер, или ранг. В том случае, если встречаются связки (т.е. повторы), им присваивается средний ранг. Например, если в выборке встретилось два значения, которые занимают ранги 9 и 10, то им обеим присваивается ранг 9,5.
Далее, для каждого наблюдаемого значения выборки рассчитывается ожидаемое нормальное значение zj по формуле:
|
|
(1) |
где N – объем выборки; Ф-1 – функция, обратная интегральной функции нормального (стандартного) распределения.
Таблицу значений Ф-1 можно найти, например, в книге Л.Н. Большева, Н.В. Смирнова (1983). Кроме того, значения zj можно получить, используя функцию НОРМСТОБР(p) табличного редактора MS Excel, где р – величина, стоящая в правой части формулы 1.
Например, для самого меньшего значения нашей выборки x[j=1] = 9,20, соответствующее значение z1 будет равно:
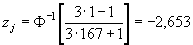 ,
,
где 167 – это объем выборки. Для максимального значения (x[j=167] = 36,70) эта величина составит, соответственно, z1 = 2,653.
Дальнейшая часть работы заключается в статистически достоверном распределении всей совокупности на группы, характеризующиеся сходным уровнем наклона (в нашем случае, это – когорты). Такое распределение может быть произведено с использованием процедуры кластерного анализа (методом k-средних) на основании двух показателей — собственно размеров тела (т.е. наблюдаемых значений) и теоретически ожидаемых значений нормальной кривой, которые были рассчитаны нами выше. Число когорт (т.е. значение k) подбирается так, чтобы кластеры характеризовались наибольшей гомогенностью. В нашем случае, на рис. 2 и 3 видно, что таких групп должно быть три.
При проведении кластерного анализа методом k-средних одной из форм вывода результатов есть таблица, где каждому исходному значению присвоен соответствующий номер кластера, к которому оно может быть отнесено. Используем эту таблицу и рассчитаем основные статистические характеристики выделенных кластеров (т.е. когорт) (табл. 1). График возрастного распределение для исходной выборки уже с учетом отнесения каждой особи к своей размерно-возрастной группе представлен на рис. 4. Отчетливо видно, что для каждой выделенной когорты характерно значимое соответствие распределения нормальному.
Таблица 1
|
Когорта |
Объем |
Размах |
Среднее арифметическое |
Стандартное отклонение |
|
1-ая |
49 |
9,2 – 16,5 |
12,15 |
1,60 |
|
2-ая |
70 |
16,8 – 26,3 |
21,05> |
2,46 |
|
3-я |
48 |
26,8 – 36,7 |
31,86 |
2,37 |
Крайние (минимальное и максимальное) значения размеров особей из каждой когорты не пересекаются. Кроме того, средние значения каждой когорты отделены от соседних не менее, чем на 2,5-3,0 стандартных отклонения (σ), т.е. отчетливо проявляется правило “трех сигм”.
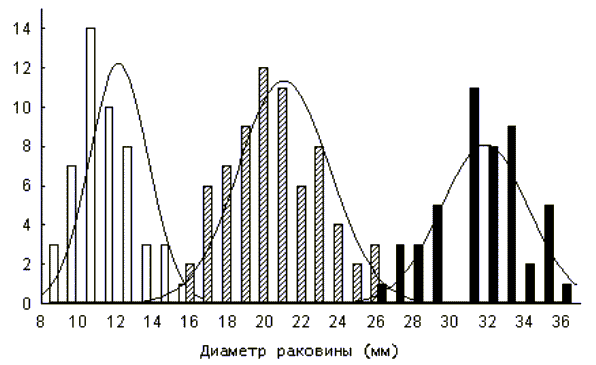
Рис. 4
Таким образом, можно считать, что первая когорта представляет собой сеголеток, вторая – годовиков, а третья – половозрелых особей возрастом 2 года и более. Поскольку размеры половозрелых особей разного возраста практически не изменяются, то разделить более дробно по возрастам их трудно.
Как и следовало ожидать, с увеличением возраста внутрикогортная изменчивость возрастает, а абсолютная частота модального класса – снижается. Это связано с тем, что численность особей старших возрастных групп снижается, а уровень их генетического (морфологического, физиологического и т.д.) разнообразия, напротив, возрастает.
2. Оценка возрастного разнообразия
Характер возрастного разнообразия популяции, можно охарактеризовать математически с использованием показателя возрастной гетерогенности (Ñ):
|
|
(2) |
где Pi — доля особей i-той возрастной группы.
Как легко убедиться, показатель возрастной гетерогенности Ñ ® 1, если популяция представлена одновозрастными особями, а для популяции с большим числом возрастных групп, представленных одинаковыми долями, Ñ ® +¥ .
Уровень возрастного разнообразия отражает реакцию популяции на пресс отбора, разная направленность и интенсивность которого во многом определяет возрастную структуру. Высокое разнообразие способствует стабильности популяции, поскольку различные стадии жизненного цикла обладают различной устойчивостью к действию экологических факторов. В крайне нестабильных, маргинальных условиях существования (а также при сильном антропогенном стрессе) возрастное распределение может значительно упрощаться.
3. Таблицы выживания и особенности их построения
Изменение численности всей популяции в целом, а также ее отдельных возрастных групп, сопряжено с двумя основными процессами — гибелью отдельных особей, находящихся на различных стадиях онтогенеза, с одной стороны, и рождением новых членов, с другой. Кроме того, на изменение соотношения различных возрастных классов оказывает влияние и процесс роста, т.е. переход особей из младших возрастных классов в более старшие. При этом растут или погибают особи не зависимо от их возраста, а вот репродуцируют только половозрелые особи (по крайней мере, те, что встретили полового партнера).
Если разбить всю популяцию на возрастные группы одинаковой продолжительности (например, один год), то численность особей любого возрастного класса (Ni), за исключением новорожденных особей, тогда будет составлять:
|
|
(3) |
где Ni-1 — численность особей предыдущего возрастного класса, а li — выживаемость особей в промежуток времени от i-1 до i (i ³ 1).
Численность новорожденных особей (N0) — нулевой возрастной группы — представляет собой сумму потомков, произведенных половозрелыми особями за единичный отрезок времени (год, в нашем случае):
|
|
(4) |
где mt, mt+1,..., mt+k — показатели повозрастной плодовитости (половозрелые особи входят в возрастные группы от t до t+k и их численности составляют, соответственно, Nt , Nt+1 ,..., Nt+k особей).
Зная значения показателей выживаемости и плодовитости, зависящих от возраста, можно более подробно проанализировать возрастную структуру популяции, динамику численности как отдельных возрастных групп, так и всей популяции в целом, рассчитать удельную скорость роста численности популяции (r) и многие другие характеристики.
Наиболее часто такой анализ проводят для научно-обоснованной борьбы с видами-вредителями, расчетов оценки запасов и уровня добычи промысловых видов, разработке программ сбережения редких и исчезающих видов.
Методика расчетов основных демографических параметров популяции при построении таблиц выживания (или демографических таблиц) описана в ряде изданий (Бигон и др., 1989; Коли, 1979; Одум, 1986; Паевский, 1985; Пианка, 1981; Солбриг, Солбриг, 1982; Федоров, Гильманов, 1980; и др.). Мы продемонстрируем эти методики на примере, приведенном ниже.
П р и м е р. Проводился учет численности особей одной когорты от момента их рождения (столбец fx в таблице 1) до возраста 9 лет (более 9 лет не прожила ни одна особь).
Таблица 1
|
Воз-раст |
Число выжив-ших |
Число погибших |
Выжива-емость |
Смерт-ность |
Удельная смерт-ность |
Удельная выжива-емость |
Интенсив-ность |
|
x |
fx |
Dx |
lx |
dx |
qx |
px |
kx |
|
0 |
472 |
251 |
1,00 |
0,53 |
0,53 |
0,47 |
0,33 |
|
1 |
221 |
55 |
0,47 |
0,12 |
0,26 |
0,74 |
0,12 |
|
2 |
166 |
37 |
0,35 |
0,08 |
0,23 |
0,77 |
0,11 |
|
3 |
129 |
30 |
0,27 |
0,06 |
0,22 |
0,78 |
0,11 |
|
4 |
99 |
24 |
0,21 |
0,05 |
0,24 |
0,76 |
0,12 |
|
5 |
75 |
21 |
0,16 |
0,05 |
0,31 |
0,69 |
0,14 |
|
6 |
54 |
19 |
0,11 |
0,04 |
0,36 |
0,64 |
0,19 |
|
7 |
35 |
17 |
0,07 |
0,03 |
0,43 |
0,57 |
0,29 |
|
8 |
18 |
16 |
0,04 |
0,04 |
1,00 |
0,00 |
0,95 |
|
9 |
2 |
2 |
0,00 |
— |
— |
— |
— |
|
Сумма |
2,68 |
1,00 |
В столбце Dx приведено количество особей, погибших между возрастом x и x+1; значения Dx рассчитываются как разность численности особей, выживших в начале возрастного класса x и в начале возрастного класса x+1:
|
|
(5) |
Например, D0 = 472 - 221 = 251; D1 = 221 - 166 = 55 и т.д.
Затем рассчитываются показатели выживаемости (lx) для каждого возрастного класса x. Данный показатель показывает, какая часть когорты доживает до конца возрастного интервала и определяет оценку вероятности новорожденной особи дожить до определенного возраста. Для новорожденных особей этот показатель, естественно, равен единице. Для остальных возрастных групп определяется как отношение числа особей, доживших до возраста x к числу новорожденных:
|
|
(6) |
Например, в нашем примере выживаемость сеголеток составляет: l1 = 221 : 472 = 0,47; двухлеток: l2 = 166 : 472 = 0,35; и т.д.
Как видно из таблицы 1, за первый год жизни выживает около половины новорожденных особей, а к возрасту 9 лет практически все погибают (выживаемость составляет 0,004 или 0,4 %).
Если на график нанести значения функции выживания организмов в зависимости от их возраста (от момента рождения до смерти), то получим график кривой выживаемости (рис. 5). Выделяют три основных типа кривых выживаемости. Тип I характерен для животных, которые в большинстве своем доживают до старших возрастных групп и затем быстро погибают. Тип II характерен для животных, вероятность погибнуть у которых в любом возрасте практически одинакова. И, наконец, тип III характерен для животных, большая часть которых погибает на ювенильных стадиях.
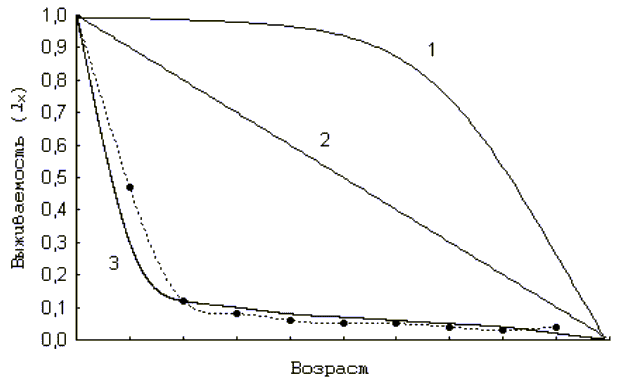
Рис. 5
Следующим показателем, который рассчитывается при построении таблиц выживания является смертность (dx). Он показывает, какая часть когорты погибнет в течение каждой из возрастных стадий и определяет вероятность особи умереть в возрастном интервале от x до x+1.
Рассчитывается показатель смертности как разность показателей выживаемости для двух последовательных возрастных классов:
|
|
(7) |
Например, смертность сеголеток на 1-ом году жизни составляет: d0 = 1,00 - 0,47 = 0,53; смертность годовиков на 2-ом году жизни: d1 = 0,47 - 0,35 = 0,12; и т.д.
Показатель смертности можно складывать при объединении возрастных классов в более укрупненные группы. Например, для особей младше 4 лет, показатель смертности составляет:
d0-3 = 0,53 + 0,12 + 0,08 + 0,06 = 0,79.
Т.е. практически 80 % особей погибает в возрасте, младше 4 лет. Естественно, сумма показателей смертности для всех возрастных классов равна единице:
|
|
(8) |
Наиболее уязвимым местом при использовании данного показателя является его низкая достоверность при маленьких объемах выборки. Более устойчив к данному ограничению показатель удельной смертности (qx). Он определяет, какая доля особей возраста x погибнет, при достижению возраста x+1 от начальной численности данной возрастной группы, т.е. определяет вероятность того, что в течение соответствующей возрастной стадии особь погибнет. Для расчета данного показателя можно использовать несколько способов:
|
|
(9) |
Чаще всего используется последнее выражение — отношение смертности к выживаемости. Например, для сеголеток удельная смертность равна: q0 = 0,53 / 1,00 = 0,53; для годовиков q1 = 0,12 / 0,47 = 0,26; и т.д.
В нашем примере удельная смертность высока для особей самых младших возрастных классов и самых старших (для последнего возрастного класса она всегда равна 1,00).
В отличие от смертности, удельная смертность более четкий показатель демографической картины и часто используется при межпопуляционных сравнениях в пределах одного вида, или при сравнении стратегии жизненного цикла близких видов.
Данный показатель нельзя складывать при объединении возрастных классов.
Кроме показателей смертности и удельной смертности (которые рассчитываются для особей каждой возрастной группы в отдельности) можно для всей популяции в целом рассчитать среднюю ежегодную смертность (d*) и среднюю удельную взвешенную смертность (q*):
|
|
(10) |
|
|
(11) |
Для этого вначале рассчитываем промежуточные значения:
S(xdx) = (0 ×
0,53) + (1 ×
0,12) + ... (8 ×
0,04) = 1,68;
S(lx ) = 1,00 + 0,47 + 0,35 + ... + 0,04 = 2,68.
Тогда, средняя ежегодная смертность будет составлять:
d* = 1 : 1,68 = 0,60, а средняя удельная взвешенная: q* = 1 : 2,68 = 0,37.
Средняя удельная взвешенная смертность показывает, какова в среднем удельная смертность для особей различных когорт. Таким образом, в среднем 37 % особей каждого возрастного класса не доживает до следующего возраста. Показатель средней ежегодной смертности близок уровню смертности для новорожденных особей (d0 = 0,53) и будет использован в дальнейшем.
Величина, обратная удельной смертности называется удельной выживаемостью (px):
|
|
(12) |
Удельная выживаемость определяет вероятность животных возраста x дожить до возраста x+1.
Наконец, используется еще один показатель, характеризующий интенсивность смертности (kx). Он рассчитывается, как десятичный логарифм отношения численности особей возраста x к численности особей возраста x+1:
|
|
(13) |
Например, для сеголеток удельная выживаемость равна: p0 = 1,0 - 0,53 = 0,47; для двухлеток: p1 = 1,0 - 0,26 = 0,74; и т.д.
А интенсивность смертности, соответственно: k0 = lg (472 / 221) = 0,33; k1 = lg (221 / 166) = 0,12; и т.д.
Вторая часть работы посвящена анализу репродуктивных особенностей популяции, оценкам продолжительности жизни особей разного возраста, средней продолжительности генерации и скорости роста численности. Пример расчета всех необходимых промежуточных величин приведен в таблице 2.
Таблица 2
|
Воз-раст |
Выжива-емость |
Удельная |
Ожидаемая |
Репродуктив-ная ценность |
||
|
x |
lx |
mx |
lxmx |
xlxmx |
ex |
vx |
|
0 |
1,00 |
0 |
0 |
0 |
2,68 |
1,13 |
|
1 |
0,47 |
0,19 |
0,089 |
0,089 |
3,57 |
2,40 |
|
2 |
0,35 |
0,35 |
0,123 |
0,245 |
3,46 |
2,97 |
|
3 |
0,27 |
0,68 |
0,184 |
0,552 |
3,19 |
3,39 |
|
4 |
0,21 |
0,62 |
0,130 |
0,520 |
2,81 |
3,49 |
|
5 |
0,16 |
2,04 |
0,326 |
1,630 |
2,38 |
3,76 |
|
6 |
0,11 |
2,21 |
0,243 |
1,458 |
2,00 |
2,51 |
|
7 |
0,07 |
0,47 |
0,033 |
0,231 |
1,57 |
0,47 |
|
8 |
0,04 |
0 |
0 |
0 |
1,00 |
0 |
|
9 |
0,00 |
0 |
0 |
0 |
0 |
0 |
|
Сумма |
1,128 |
4,725 |
Наиболее трудоемкой частью подготовительного этапа для расчета параметров, представленных в таблице 2 является оценка удельной плодовитости животных различных возрастных групп (mx). Он дает оценку среднего числа новорожденных потомков, произведенных одной особью в интервале между возрастом x и x+1. Также он может быть выражен числом новорожденных самок на одну самку того или иного возраста.
Для млекопитающих для расчета удельной плодовитости может быть использована формула:
|
mx = f· g· h· v, |
(14) |
где f — среднее число эмбрионов на одну самку за одну беременность; g — доля размножающихся самок среди всех самок; h — доля самок в общей численности популяции; v — число пометов на одну самку за год.
Для яйцекладущих животных:
|
mx = k· n· p· s1 ·s2, |
(15) |
где k — среднее число кладок за год; n — средний объем кладки; p — доля животных, откладывающих кладки; s1 — уровень вылупляемости; s2 — выживаемость новорожденных сразу после вылупления.
В нашем примере, плодовитость нарастает со второго года жизни, достигая максимума (примерно 2,2 новорожденные самки на 1 самку) в возрасте 5 лет, а затем вновь снижается (до 0 у 8-9-летних особей). Показатели удельной смертности имеют практически противоположную зависимость (табл. 1): за исключением сеголеток, удельная смертность особей младших возрастных групп очень низка, а в возрасте 5-6 лет она заметно повышается. Таким образом, можно предположить, что повышение энергетических затрат на репродукцию резко снижают выживаемость особей.
Средняя по возрастам удельная плодовитость представляет собой среднее арифметической средней плодовитости среди животных возрастных групп, среди которых отмечается репродукция. В нашем случае этот показатель составляет: (0,19 + 0,35 + ... + 0,47) : 7 = 0,94. Другими словами, в среднем, каждая самка, начиная со второго года жизни, приносит только по одному детенышу женского пола.
Другим показателем, характеризующим способность реализации репродуктивного потенциала особями изучаемой популяции, является чистая скорость размножения (R0). Этот показатель определяется как среднее число потомков нулевого возраста, произведенных организмом за всю его жизнь.
Рассчитывается данный показатель, как сумма произведений специфической для каждого возрастного класса выживаемости и удельной плодовитости:
|
|
(16) |
В нашем примере, чистая скорость размножения (или коэффициент размножения на одно поколение) составила 1,13, т.е. в среднем каждая самка в течение всей ее жизни оставляет 1,13 особей женского пола.
Ожидаемая продолжительность жизни (ex) особей разного возраста оценивает, сколько в среднем еще проживет особь, достигшая возраста x. Для любого возраста, ex — это средняя продолжительность оставшейся жизни особей.
Данный показатель оценивается по формуле:
|
|
(17) |
Например, ожидаемая продолжительность жизни для сеголеток составляет:
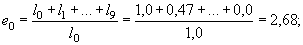
для годовиков:
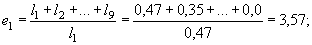
и т.д.
Таким образом, для новорожденных особей средняя продолжительность жизни составляет 2,68 лет; если особь прожила первый год своей жизни, она в среднем проживет еще 3,57 лет, и т.д. Как видно из таблицы 2, максимальная ожидаемая продолжительность жизни имеет место для особей, проживших первые год или два своей жизни, а затем она постепенно снижается. Для восьмилеток, как и следовало ожидать, срок будущей жизни в среднем равен одному году.
Общая средняя продолжительность жизни для животных всех когорт рассчитывается по формуле:
|
|
(18) |
где d* — средняя ежегодная смертность (см. выше). Таким образом, для животных из рассматриваемого примера общая средняя продолжительность жизни составляет чуть более одного года:
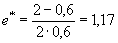 .
.
Еще одним временным популяционным показателем является среднее время генерации (Tc). Среднее время генерации, или средняя продолжительность поколения, представляет собой средний возраст всех самок в популяции, взвешенный в соответствии с производимым им потомством.
Данный показатель рассчитывается по формуле:
|
|
(19) |
Для рассматриваемого примера, среднее время генерации составляет чуть более 4 лет: Tc = 4,725 : 1,128 = 4,19.
Именно для животных, проживших четыре первые года своей жизни, удельная плодовитость резко повышается. В этом возрасте происходит половое созревание большей части когорты и они вступают в размножение. Как будет показано ниже, среднее время генерации в значительной мере определяет скорость роста численности популяции и многие популяционные характеристики (например, репродуктивную стратегию).
Следующей характеристикой популяционной структуры является показатели репродуктивной ценности (vx) для особей разных возрастных групп. Для особей возраста x, репродуктивная ценность определяется как специфическое для этого возраста ожидание будущего потомства. Математически данная величина рассчитывается по формуле:
|
|
(20) |
Для годовиков репродуктивная ценность составляет:
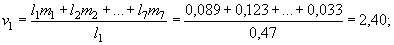
для двухлеток:
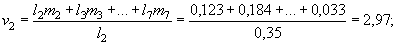
и т.д.
Для новорожденных особей показатель репродуктивной ценности, как и следовало ожидать, равен чистой скорости размножения, т.е. v0 = R0; а для последнего возрастного класса, представители которого еще способны к репродукции — специфической для него удельной плодовитости (т.е. в нашем примере v7 = m7).
Наконец, зная чистую скорость размножения (R0) и среднее время генерации (Tc) можно рассчитать удельную скорость роста популяции (r). Данный показатель имеет очень большую важность, поскольку практически однозначно характеризуют судьбу популяции: если r > 0, численность такой популяции возрастает; если r < 0, напротив, численность такой популяции постепенно снижается; и, наконец, если r = 0, то такая популяция находится в стационарном состоянии.
Значение скорости роста популяции приближенно можно получить по формуле:
|
|
(21) |
Как видим, определяющим для получаемого значения является чистая скорость размножения, т.е. среднее число потомков произведенных особью за всю свою жизнь. Если этот показатель равен единице, каждая погибшая особь оставляет после себя 1 потомка, численность популяции остается постоянной и r = 0. Если же R0 больше или меньше 1, численность популяции нарастает или снижается, соответственно.
В нашем примере, скорость роста популяции равна: 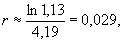 т.е. очень близка к нулю.
т.е. очень близка к нулю.
П.Лесли (1945) показал, что скорость роста популяции может быть найдена как корень следующего уравнения:
|
|
(22) |
где l = er; а n — число возрастных групп. Другие обозначения приведены выше.
Для нашего примера, учитывая, что удельная плодовитость новорожденных, а также 8-ми и 9-ти летних особей равна нулю, получаем уравнение:
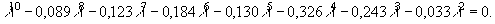
Данное уравнение трудно решить алгебраически, однако, применив метод итераций, можно с определенной точностью определить значение l, при котором выражение слева превращается в ноль. В данном случае это значение равно 1,023.
Следовательно, скорость роста популяции:
r = ln 1,023 = 0,023.
Как видим, значения r, полученные двумя разными способами, оказываются очень близки.
Кроме того, оценка l дает возможность рассчитать, во сколько раз увеличится численность популяции за один год.
Эту величину можно еще рассчитать по формуле:
|
|
(23) |
* * * * *
Наиболее важной задачей при составлении демографических таблиц является получение с достаточным уровнем значимости величин fx. Существует четыре основных метода их получения (Коли, 1979).
1. Прослеживают судьбу когорты от момента рождения особей до гибели последнего ее представителя. Через равные промежутки времени регистрируют число выживших членов. Эти значения и будут оценками fx. Параллельно может вестись учет нескольких когорт, рожденных через равные промежутки времени. В итоге все полученные значения уровня выживаемости для каждой отдельной возрастной группы суммируют. Данный метод подразумевает мечение особей (индивидуальное или групповое; представителей одной когорты — одной меткой, а представителей разных — разными). Методы оценки численности, которые подразумевают мечение особей в популяции приведены во второй части Практикума.
2. Если после гибели особи остается долго хранящиеся останки (например, костные — черепа, рога, зубы и т.п.) по котором можно точно определить возраст погибшего животного, то, учитывая число особей погибших в каждом возрасте (т.е. показатели Dx; см. таблицу 1), можно рассчитать соответствующие значения fx следующим образом:
f7 = D7;
f6 = D7 + D6;
f5 = D7 + D6 + D5;
f4 = D7 + D6 + D5 + D4; и т.д.
Чем больше используется материальных остатков и чем надежнее определен возраст, тем достовернее получаемые демографические показатели.
3. Если известным способом оценить возрастной состав особей исследуемой популяции в момент времени i, а затем провести такую же операцию в момент времени i+1, то разность между численностью особей возраста x в первой выборке и численностью особей возраста x+1 во второй даст оценку числа погибших особей (т.е. Dx). Кроме того, если эти полученные показатели разделить на исходную численность особей возраста x, то можно сразу получить оценки удельной смертности (qx), на основании которых легко рассчитываются показатели выживаемости (lx) и смертности (dx).
П р и м е р. В таблице 3 приведены значения численности (Nx) особей различного возраста в момент времени x и x+1.
Таблица 3
|
В о з р а с т (x) |
||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
|
|
Ni |
300 |
250 |
150 |
50 |
20 |
10 |
|
Ni+1 |
250 |
150 |
100 |
70 |
30 |
5 |
Рассчитаем значения удельной смертности:
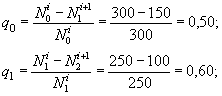
и т.д.
Зная, что выживаемость нулевого возрастного класса всегда равна единице (т.е. l0 = 1,00), смертность для особей этого возраста, следовательно, равна удельной смертности (т.е. d0 = q0). Для всех последующих возрастных групп показатели выживаемости и смертности рассчитываются по индукции:
|
|
(24) |
Например, для годовиков выживаемость равна: l1 = l0 - d0 = 1,00 - 0,50 = 0,50; а смертность: d1 = l1·q1 = 0,50·0,60 = 0,30, и т.д.
В итоге получаем полностью заполненную таблицу (табл. 4), и используя показатели удельной плодовитости (которые определяются постановкой отдельных природных или природно-лабораторных экспериментов) можно рассчитать все остальные демографические показатели.
Таблица 4
|
x |
lx |
dx |
qx |
|
0 |
1,00 |
0,50 |
0,50 |
|
1 |
0,50 |
0,30 |
0,60 |
|
2 |
0,20 |
0,11 |
0,53 |
|
3 |
0,09 |
0,04 |
0,40 |
|
4 |
0,05 |
0,04 |
0,75 |
|
5 |
0,01 |
0,01 |
1,00 |
Основным ограничением использования данного метода является уровень точности оценок численности и равномерность отлова особей различного возраста во время двух последовательных учетов.
4. Наконец, если выборочное возрастное распределение за несколько последовательных лет исследования не проявляет резких изменений соотношения численности особей разного возраста, то последние показатели могут служить (в первом приближении) оценками fx.
В этом случае относительная стабильность возрастного распределения свидетельствует о низкой вариабельности специфических для каждой возрастной группы показателей выживаемости и удельной плодовитости. Стабильность возрастного распределения может быть оценено, используя например критерий Хи-квадрат Пирсона для сравнения частот нескольких выборок или его модификации.
Однако может случиться так, что численность особей возраста x+1 будет больше численности особей возраста x (случай т.н. “отрицательной смертности”). В такой ситуации выборочное возрастное распределение подвергают сглаживанию.
Можно использовать различные модели регрессии для сглаживания эмпирических распределений, подбирая при этом самую оптимальную (уровень оптимальности оценивается на основе величины коэффициента детерминации — R2. Он определяет, какая доля изменчивости исходного набора данных описывается используемой моделью. Желательно, чтобы R2 ³ 90 %).
Например, часто используется для расчета теоретических значений fxтео полиномная модель:
|
|
(26) |
или логарифмически-полиномная:
|
|
(27) |
где x — возраст организма. Необходимо, чтобы на рассматриваемом отрезке временнó й оси (ось OX) проходила нисходящая ветвь линии регрессии (для данных функций характерно наличие как нисходящих, так и восходящих ветвей).
Часто используют функции типа:
|
|
(28) |
или различные ее модификации. (Поскольку первый возрастной класс имеет возраст 0, а на 0 делить нельзя, то к знаменателю формулы прибавляют единицу).
Можно также использовать функцию:
|
|
(29) |
(При некотором возрасте линия регрессии пересекает ось OX, и получаются отрицательные значения теоретических частот, т.е. необходимо проверять, чтобы в пределах рассматриваемого отрезка времени этого не происходило).
Наконец, можно избежать “отрицательной смертности”, используя метод линейного сглаживания по трем ординатам:
|
|
(30) (31) (32) |
где n — номер последнего возрастного класса.
Г.Коли (1979) также предлагает для сглаживания эмпирического возрастного распределения использовать пробит-функцию:
|
|
(33) |
где N — суммарная численность особей всех возрастных групп (т.е. N = S fx).
Пробит-функция имеет вид:
|
|
(34) |
где Ф-1 – функция, обратная интегральной функции нормального (стандартного) распределения (см. формулу 1).
Литература
| Выпуск 2
|
Начало |
Список
|
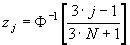 ,
,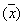
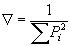 .
.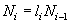 ,
,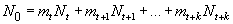 ,
,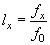 .
.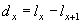 .
.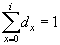 .
.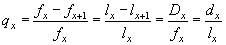 .
.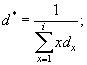
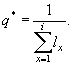
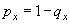 .
.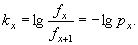
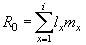 .
.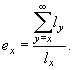
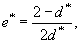
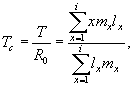
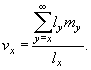
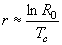 .
.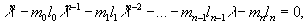
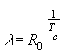 .
.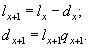
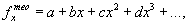
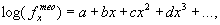
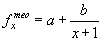
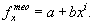
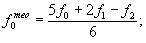
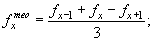
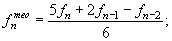
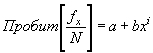 ,
,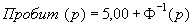 ,
,