© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Опубликовано: Морской экологический журнал (Marine ecological Journal).- 2006.- Том V, № 3. - С. 31-44.
УДК 574.9+577
БИОМЫ В ГИДРОСФЕРЕ
А.А.Протасов, докт. биол. наук, вед. научн. сотр.
Институт гидробиологии Национальной Академии наук Украины, Киев, Украина
Ключевые слова: биом, гидросфера, биосфера Земли, гидробиология, биогеография гидросферы.
Эпиграфом данной статьи вполне может быть замечание, принадлежащее болгарским гидробиологам:
Гидроэкосистемы не есть просто “мокрый вариант” хорошо изученных наземных биомов и биогеоценозов.
Узунов Й., Ковачев С., [46, с. 239]
Сравнительный анализ водных и наземных экосистем проводился как в биогеографическом [20, 21], так и в гидробиологическом аспектах [22, 12, 29]. Качественные и количественные различия факторов среды для гидро- и атмобионтов велики и достаточно очевидны. Например, Л.А.Зенкевич [21] среди физических и химических факторов выделяет “типичные, особенно для больших глубин океана, однообразие и устойчивость факторов среды на больших пространствах”. Условия среды значительно изменяют пути вещества, энергии и информации в экосистемах [2]. Например, как было отмечено Г.Г.Винбергом, “большая часть океана, а, следовательно и большая часть нашей планеты занята сообществом гетеротрофных организмов, нуждающихся в притоке органического вещества” [12 (с. 538)], в то время, как на суше наблюдается подавляющее преобладание автотрофных. Условия в гидросфере и на суше (среда обитания атмобионтов) существенно различаются, поэтому биотическую часть экосистем составляют совершенно различные жизненные формы, экоморфы. При этом общие закономерности функционирования водных и наземных экосистем остаются принципиально сходными.
Положения одного из системных обобщений, а именно – функционально сходные системы могут состоять из различных элементов [42] – предполагают, что явление экологической конвергенции должно иметь широкое распространение. Подчеркивая различия видового состава в тех или иных географических областях, Ю.Одум [38] указывал на то, что сходство физической среды определяет развитие сходных экосистем. Как отмечал П. Джиллер [17], "существование различных типов местообитаний, “которые можно сгруппировать в биомы, подразумевает некую форму конвергенции сообществ” (с. 29). Нами [41] было предложено использование принципов биоценотической конвергенции и биоценотического градиента в анализе различных по видовому составу, но близких по составу экоморф сообществ гидробионтов.
Еще в 1807 А. Гумбольдт (A. Humboldt) г, указывал (цит. по [24]): "Как отдельным органическим существам присуща своя физиономия, так существует и физиономия природы, принадлежащая исключительно одному климату" (с. 491). То есть, сходство условий формирует физиономическое сходство сообществ. На суше это физиономическое сходство определяется в первую очередь сходством облика растительности. По Гумбольдту около 20 жизненных форм растений определяет “в основном физиономию природы” [24]. Концепция жизненной формы, экоморфы, имеет достаточно богатую историю и библиографию [1, 23, 24, 27, 34]. Важно то, что биотические сообщества при всем их многообразии имеют существенные черты сходства и это сходство в значительной мере определяется сходством ключевых для этих сообществ жизненных форм. Многие геоботанические и экологические данные и обобщения [11, 38] позволяют вывести общую закономерность физиономического сходства растительности, набора жизненных форм, которые характерны для тех или иных условий. Эту общую закономерность можно определить как принцип сходного распространения близких жизненных форм или закон соответствия жизненных форм условиям среды А. Гумбольдта. Как писал Д. Н. Кашкаров, [27] “в жизненной форме”, как в зеркале, отражаются главные черты местообитания животного” (с. 121). В самом широком смысле этот закон можно применить в разделении атмобионтов и гидробионтов как наиболее общих жизненных форм в соответствии с их средой обитания. Узко приспособленные формы могут индицировать более дробные особенности местообитаний.
Идея соответствия состава организмов условиям их обитания имеет достаточно давнюю историю. Чарльз Лайель (С. Lyell), цит. [25] отмечал, что еще в конце XVIII в. Ж.-Л. Бюффоном был сформулирован общий закон в географическом распределении организмов, согласно которому географические регионы населены различными видами животных и растений.
В плане исторического взгляда на развитие экологической мысли важно отметить, что Ч. Лайель указывал: “Бюффон… допускает, что следовало бы ожидать, что равная температура и другие одинаковые условия существования должны были производить одинаковые существа” (цит. по [25], (с. 25), то есть речь шла о принципе соответствия состава сообществ условиям обитания задолго до рождения экологии Э. Геккеля.
Экологические и биогеографические наблюдения, таким образом, приводят к представлениям об определенных закономерностях районирования и распределения организмов по поверхности Земли, а также к представлениям об определенном распределении и сходстве не только видового состава населения, но и жизненных форм.
В основе географического районирования лежит понятие географического ландшафта. Он представляет собой совокупность особенностей рельефа, климата, вод, почвенного и растительного покрова, животного мира, другие особенности, включая результаты деятельности живых организмов и человека, которые “сливаются в гармоничное целое, типически повторяющиеся на протяжении данной зоны Земли” [7], (с. 5). Ландшафт рассматривался Л. С. Бергом как “географический индивид” или “особь”.
Продолжая эту аналогию с особями можно задаться вопросом: что представляет собой популяция таких “особей”? Аналогия, безусловно, весьма поверхностная, однако можно полагать, что в составе такой “популяции” будут находиться ландшафты, обладающие определенной индивидуальностью, то есть в популяции в целом – разнообразием, но, в то же время, они будут сходными, поскольку вошли в состав “популяции”. Далее – совокупность таких популяций занимает некий ландшафтный “ареал” - по аналогии с видовым ареалом.
Видовой ареал, как известно, представляет собой часть поверхности Земли, в пределах которого обитает тот или иной вид организмов [32]. Таким образом, под “ареалом” ландшафтов мы можем понимать область (часть) земной поверхности, занятую теми или иными сходными, однотипными ландшафтами. Ландшафтные “ареалы” также как видовые могут быть и дизъюнктивными.
При проведении дальнейшей аналогии в системном плане представляется вполне логичным сравнением ландшафта с экосистемой. При этом следует учесть важное замечание А. Н. Миронова [36] о том, что биологические системы относятся к группе совмещаемых в пространстве, в том случае если их ареалы (ареалы популяций и таксонов) накладываются друг на друга. Это касается, очевидно, именно совмещения ареалов, так как ни совмещения в пространстве отдельных организмов, ни сообществ невозможно. Также невозможно совмещение в пространстве ландшафтов, биокосных, но геоцентрических систем.
Видимо, можно согласиться с К. М. Петровым [40], что различия между ландшафтом и экосистемой обусловлено лишь смещением акцентов. (Не совсем ясно, однако, почему, А. Н. Миронов [36] эту точку зрения связывает только с подводным ландшафтоведением).
Мы рассматриваем экосистему вслед за А. Ф. Алимовым [2] как локализованную в пространстве и динамичную во времени совокупность совместно обитающих и входящих в сообщества различных организмов и условий их существования, находящихся в закономерной связи друг с другом.
Взгляд на ландшафт геоцентричен, на экосистему – биоцентричен (рис. 1). Экосистема функционирует, что обусловлено динамическим взаимодействием потоков вещества, энергии и информации. Ландшафт изменяется, трансформируется, и во многом – за счет биотических процессов.
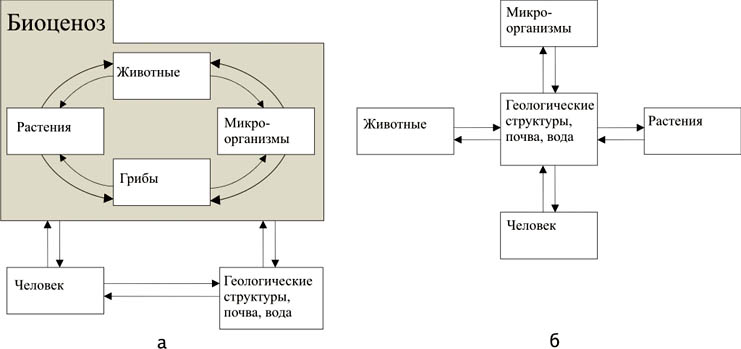
Рис. 1. Структура экосистемы (а) и структура ландшафта (б)
[Fig.1. The structure of ecosystem (a) and structure of landscape (b)]
Относительное сходство между ландшафтом и экосистемой не бесспорно. Мы усматриваем, однако, полезность такого параллелизма в том, что эти два понятия должны дополнять друг друга. С легкой руки Ю.Одума понятие “экосистема” практически потеряло свое хорологическое значение, став почти полностью функциональным [2, 38]. Именно необходимость пространственного взгляда на структуру геомериды заставляет прибегать к понятию и термину “биогеоценоз”, подчеркивая его хорологичность [24, 43].
Распределение живых организмов, в соответствие с законом Бюффона имеет определенную зональность, В. В. Докучаев в конце XIX в распространил учение о зональности на весь земной шар [7], как на почвы, так и на весь природный комплекс, включая человека и различал от высоких широт к низким бореальную или тундровую зону, таежную или лесную зону, черноземную или степную, а также зону сухих субтропических стран. Климат ландшафтных зон разнообразен и неоднороден. Например, на северо-западном побережье Европы океаническо-арктический климат с умеренно холодной зимой и прохладным летом. На севере Сибири климат континентально арктический с суровой зимой и относительно теплым летом [11]. Однако, анализ климадиаграмм во многих пунктах Арктической зоны показывают принципиально сходную картину [11, 13]: существенные годовые сезонные колебания температуры ( 30 и более °С) и относительно высокое количество осадков летом. Полярная древесная растительность распространяется на север приблизительно до изотермы июля +10 °С. В зоне тропических дождевых лесов, напротив – колебания высокой (25-28єС) температуры в течение года минимальны, количество осадков велико и имеет несколько пиков.
В соответствии с климатическими условиями формируется определенный тип растительной формации, а также вся биота данной климатической зоны. Важно отметить, что ареалы наземных ландшафтов, также как и ареалы видов имеют вполне определенные границы. Они образует структуру витасферы (термин Тюрюканова, Александровой, [47]), или живой части биосферы Земли, точнее – части витасферы, связанной с материками.
Принцип зонального соответствия условий в том или ином регионе и населяющей его биоты обобщен в понятии биома: “региональный климат, взаимодействуя с региональной биотой и субстратом, образует крупные, легко выделяемые единицы сообществ, называемых биомами” [38] ( с. 489).
Как всякое обобщающее понятие, биом получил различные трактовки и интерпретации. Существуют очень широкие определения, например, И. И. Дедю [16] определяет биом (biomе) как биотическое сообщество, главную биотическую зону, макроэкосистему, совокупность сообществ. Имеются и крайне узкие определения: биом рассматривается как совокупность организмов, живущих в пределах водного объекта или региона [30, 39, 44].
Лишь в немногих определениях подчеркиваются различия наземных и водных биомов: “биом – это биогеографический регион или формация большое сообщество, которое характеризуется различными жизненными формами и основными видами растений (наземные биомы) и животных (морские)” [59], (p. 33).,
Во всяком случае, к понятию биома прибегают тогда, когда необходим общий взгляд на совокупность группировок различных живых организмов или биоценозов. Так, В.И. [19] рассматривал биом как “сложный биоценоз” или совокупность связанных биоценозов в водоемах: “биом представляет собой исторически сложившееся население водоема, состоящее из биоценозов, связанных между собой общностью места обитания” (с.62, курсив наш, А.П.). При этом следует отметить особенности используемой им системы биоценозов, а именно, выделение их на нескольких уровнях по критериям топическим (более общий подход, например, выделение литореофильных биоценозов) и по критериям биотическим (более частный, например, выделение реофильных биоценозов дрейссены). При этом В.И. Жадин подчеркивал, что “во всех случаях при наименовании биоценозов по руководящей форме следует исходить не из формально-статического вычисления преобладания одного или двух видов,… а из наличия организма, своей жизнедеятельностью накладывающего особый отпечаток на весь биоценоз” [19] с. 54). При этом им не делалось оговорки, что могут существовать и существуют биоценозы, в которых средообразующая, эдифицирующая роль какого-то определенного вида или жизненной формы не выражена. То есть в ценозе могут присутствовать, а могут и отсутствовать “организмы-инженеры экосистем” (“organisms - ecosystems engineers”) [57, 58]. Здесь также просматривается параллель с ландшафтами, одни из которых насыщены жизнью, облик их определяется биологическими компонентами (лесной ландшафт, коралловый риф), в других – облик определяется абиотической компонентой (пустыни, открытый океан).
Очевидно, что биом имеет двойственную природу как категория биогеографическая и как экологическая. Если рассматривать биом как совокупность различных групп организмов и среды их обитания в определенных широтной и вертикальной зонах [26], то эти группы можно рассматривать и как биотические комплексы [31], и как компоненты определенных ландшафтов. И то и другое определяется биогеографическим подходом. В то же время закономерно изменяющиеся условия среды определяют формирование различных по составу видов и жизненных форм, продуктивности, трофической и хорологической структуре сообществ. Так, в науке о растительности понятию "биом" соответствует понятие "растительная формация" [35], которая, следует заметить, имеет определенные физиономические черты, количественные характеристики, определяемые условиями данного региона. То есть растительность выступает маркером условий (возвращаемся к концепции А. Гумбольдта).
Таким образом, как и в случае с ландшафтами и экосистемами, мы приходим к выводу о двойственном - географическом и экологическом (геоцентрическом и биоцентрическом) подходе к одному явлению.
Сравнение распределения на поверхности материков основных биомов [11, 13, 38] и картосхемы распределения биомассы на Земле [50], а также первичной продукции [54] вполне определенно показывает их совпадение. Обобщенный подход к оценке продуктивности, который явно опирается на выделение биомов, позволяет определить несколько классов экосистем по их продуктивности [33, 49]: в тропическом дождевом лесу продукция составляет в среднем 2000-3000 г ·м-2 ·год-1; а в пустынях, полупустынях, тундре – менее 250 г· м-2 ·год-1.
Использование концепции биома – то есть обобщенного взгляда на распределение ландшафтов-экосистем с определенным осреднением их качеств и свойств позволяет более системно подходить к районированию биосферы в целом. Именно такой подход используется в глобальных характеристиках биосферных явлений [38, 49, 53]. Он позволяет делать важные экологические обобщения, подобные следующим: “Тундра – это в сущности влажный арктический злаковник” [38], с. 491) или - “огромное разнообразие видов на локальных пространствах дождевого тропического леса и в коралловых рифах общеизвестно” [52], р.1302.
Экосистемы не только отличаются своей сложностью, но являются элементами более сложных систем. Эта иерархия не только экосистем, но и биомов [40] является одной из важных составляющих структуры биосферы в целом.
Биом не всегда имеет четко установленные границы [37], однако является не меньшей биогеографической реальностью, чем видовой ареал. В экологическом смысле биом представляет собой систему экосистем, “элементарных ландшафтов”. Именно систему, а не простую сумму их, поэтому обладает определенными эмерджентными свойствами, в настоящее время недостаточно еще изученными и обобщенными. Эмерджентность не позволяет считать биом просто некоей большой экосистемой или “суперэкосистемой” в смысле простого суммирования малых. Биом должен рассматриваться как один из уровней организации биосферы.
Можно прийти к заключению, что в “наземной” экологии и биогеографии само понятие биома и его практическое использование в хорологическом анализе подразделений биосферы вполне закрепилась. В гидробиологии, морской и континентальноводной биогеографии понятие биома используется крайне мало. Это выглядит парадоксом при рассмотрении биосферных процессов в их единстве. Определение биома в гидробиологическом аспекте [26] строится на связи его с зонами – широтными и вертикальными, однако в указанной работе авторы понятие биома практически не используют, также как, впрочем, и В.И Жадин [19] в своем обзоре гидробиологических задач и проблем. В работе Л.А. Зенкевича [20] мы находим термин “биома”, который обозначает животное и растительное население вместе, служащий для обозначения “биомических комплексов”, установленных на основе количественного распределения гидробионтов. Понятие биома и здесь играет вспомогательную роль.
Из известной нам литературы, пожалуй, наиболее близки к системному обобщенному взгляду на гидробиомы болгарские исследователи И. Узунов и С. Ковачев [48]. В первую очередь следует обратить внимание, что к выделению главных, основных местообитаний они подошли чисто гидробиологически: это - плейсталь, пелагиаль и бенталь. При этом они учитывают, что “малые”, местообитания могут быть типизированы с учетом “течения и перемешивания воды, физических и химических градиентов, потоков вещества и энергии” (с.239). Далее эти авторы проводят весьма показательное сравнение между наземными и водными биомами. Например, поверхность последних относится к первым как 1: 2.5, объем, соответственно, как 1: 99, чистая первичная продукция как 1: 0.54, вторичная – 1: 3.54.
Рассматривая далее вопрос о биомах гидросферы, Узунов и Ковачев выделяют так называемые "типовые экосистемы": морские (открытый океан, континентальный шельф, зоны апвеллингов, эстуарии) и пресноводные (лентические или стоячие воды, лотические или текучие воды, заболоченные территории). Именно необходимость выделения типичных экосистем является чаще всего поводом для использования самого термина “биом” в гидробиологических исследованиях. Например, широко используется понятие "коралловый биом" [55, 56], когда необходимо представить некоторую обобщенную характеристику всех коралловых экосистем.
Мировой океан представляет собой единую оболочку Земли, существуют представления, о нём как о едином биоме [51], однако известно, что в нем дифференцируются как топические, так и биогеографические, биоценотические неоднородности [3, 4, 10, 20, 26, 40, 46]. Для континентальных вод наоборот, более или менее выраженная дискретность является одной из их характеристик. В гидросфере могут быть выделены разномасштабные хорологические и функциональные единицы – биомы, сопоставимые с наземными, однако, очевидно, что принципы их выделения отличаются от выделения последних.
Для наземных биомов, если рассматривать их как систему живого населения и определенных биотопов, формула биома может выглядеть следующим образом: климатические условия (температура, влажность) + характер почв + ключевые жизненные формы (экоморфы) организмов + трофическая структура (источники энергии, основные трофические связи) + хорологические характеристики (характер границ, ярусность, стратификация, мозаичность) или в краткой форме - КУ+П+ЖФ+Тр+Х.
Если взять в качестве примера хорошо изученные биомы суши, то в соответствие с приведенной формулой можно дать следующую характеристку некоторых из них (табл. 1)
Таблица 1. Характеристика некоторых биомов суши
Table 1. The characteristics of some land biomes
|
Биом |
Климат |
Почва |
Основные жизненные формы, |
Трофические связи |
Пространственная структура |
||||
|
температура |
Влажность |
||||||||
|
Растения |
Животные |
Первичнопро–дуценты |
Основные трофические связи |
Границы, распространение |
Ярусность, пространственная сложность |
||||
|
Степной (степи, прерии, пампа) |
Средне–годовая температура положительна, хорошо выражена сезонность температур |
Влажность умеренная, выражены сезоны засухи |
Черноземы разных типов, богатые гуматами |
Травы |
Крупные фитофаги, землерои |
Хлорофитные фотоавтотрофы |
Фотоавтотрофы - фитофаги |
Умеренная зона |
Сложность невысокая, существует микроярусность |
|
Дождевой тропический лес, гилея |
Высокие температуры без выраженных сезонных колебаний |
Влажность значительная, большая часть года увлажнение превышает испарение |
Глинистые и суглинистые, кислые, богатые окислами железа |
Древесные, лианы, эпифиты |
Древолазы, разруши-тели древесины, первичные детритофаги |
Хлорофитные фотоавтотрофы |
Фотоавтоторофыпервичные фитофаги- детритофаги |
Приэкваториальная, тропическая зона |
Сложность очень высокая, ярусность выражена |
Для гидробиомов существенное значение имеют иные, чем для биомов суши характеристики среды. Определенной спецификой обладают и сообщества гидробионтов. Климатические условия детерминируются термическим режимом и характерной повторяемостью гидродинамических условий. Если в наземных биомах всегда доминируют фотоавтотрофы и основным источником энергии является Солнце, то в гидросфере существуют и другие условия, где хемоавтотрофы играют важную роль первичнопродуцентов [8]. Кроме того, существуют гидроэкосистемы, в которых автотрофные организмы практически отсутствуют, жизненные процессы поддерживаются за счет органического вещества, производимого в других экосистемах, это глубоководные экосистемы афотической зоны [5, 12].
Рассматривая биом как совокупность сходных экосистем и ландшафтов, мы неизбежно сталкиваемся с различными градациями этого “сходства”. Сходные на одном уровне, экосистемы могут различаться на другом.
Все наземные биомы, при всех их различиях при самом общем взгляде представляют собой, по сути, лишь варианты одного эпиконтинентального гипербиома. В гидросфере исторически сложились и биотопически принципиально различаются биомы океаносферы и биомы поверхностных вод суши. Вероятно, справедливо назвать первые первичными гидробиомами, поскольку и среда обитания организмов, и собственно биота по своему происхождению прямо связаны с историей становления океана. Вторичные биомы – это биомы континентальных вод. Среда обитания организмов в них формируется как следствие взаимодействия поверхностного стока вод и литосферы, а биота имеет происхождение как из первичных гидробиомов, так и из биомов суши. Экосистемы континентальных вод тесно связаны с экосистемами суши, характер многих ландшафтов в значительной мере определяется наличием водоемов и водотоков. Тем не менее, экосистемы последних в силу своей специфики, безусловно, принадлежат к одной категории гидробиомов, как и морские.
Формула биома гидросферы выглядит следующим образом: термические условия + динамика водных масс (интенсивность водообмена, скорость и характер течений) + соотношение окислительных и восстановительных условий + субстрат + ключевые жизненные формы биоты (экоморфы, ценоэкоморфы) + трофическая структура (источники энергии, ключевые трофические связи) + хорология (границы, ярусность, стратификация, мозаичность) или Т+Д+ОВ+С+ЖФ+Тр+Х. Исходя из этой формулы могут быть представлены следующие основные характеристики гидробиомов (табл 2).
Таблица 2. Основные характеристики гидробиомов
[Table 2 . The mane characteristics of hydrobiomes]
|
Биом |
Условия среды |
Субстрат |
Основные жизненные формы |
Трофические характеристики |
Хорологическая характеристика |
||||||
|
температура |
Водообмен |
Течение |
|||||||||
|
Прокариоты |
Растения |
Животные |
Первичнопродуценты |
Основные трофические связи |
Границы, распространение |
Ярусность, пространственная сложность |
|||||
|
Пелагический океанический |
Значительная часть глубинных водных масс термически стабильна. В поверхностных слоях выражена широтная зональность температур, нарушаемая течениями. |
Очень замедленный |
Поверхностные, глубинные, вихревые и циркулярные течения, глобальный “конвейер” течений. |
Водная среда как опора |
Взвешенные в воде микроформы |
Взвешенные в воде микроформы |
Взвешенные в воде микроформы ,подвижные макроформы, активные пловцы. |
Микроскопические и субмикроскопические хлорофитные фотоавтотрофы. |
Фотоавтотрофы – первичные фаготрофы (фотичекая область). |
Весь Мировой океан, широтная зональность выражена, особенно в поверхностных слоях. |
Выражена глубинная стратификация, разномасштабная гетерогенность, определяемая динамикой водных масс. |
|
Прибрежный биом океана |
Выражена широтная зональность, нарушаемая течениями и апвеллингами. |
От слабого до интенсивного. |
Прибрежные течения, апвеллинги, даунвеллинги. |
Рыхлый и твердый субстрат разной дисперсности, водная среда как опора . |
Взвешенные в воде микроформы, микроорганизмы на субстрате. |
Взвешенные в воде микроформы, микро и макроформы на субстрате |
Взвешенные в воде микроформы . |
Микроскопические и субмикроскопические плавающие хлорофитные фотоавтотрофы. |
Разнообразные пастбищные и детритные цепи питания. |
Океанические и морские прибрежья, шельфовые зоны. |
Глубинная стратификация., зональность, связанная с приливными явлениями. Выражена топическая мозаичность . |
|
Биогермовый. |
Высокая, стабильная. |
Значительный. |
Прибрежные течения. |
Зоогенные и фитогенные карбонатные постройки |
Взвешенные в воде микроформы, микроорганизмы на субстрате |
Хлорофитные симбиотрофы, прикрепленные микро- и макроформы |
Седентарные симбиотрофы, хищники, фильтраторы |
Симбиотрофы, прикрепленные хлорофитные фотоавтотрофы |
Симбиотрофия, |
Циркумтро–пическое. |
Пространственная сложность очень высокая. |
|
Батиально-абиссальный |
Низкие и стабильные температуры. |
Замедленный |
Слабые |
Донные отложения, илы, твердые минеральные породы. |
Бактериальные пленки на субстрате. |
Отсутствуют. |
Закапывающиеся в грунт, седентарные |
Отсутствуют |
Детритные цепи питания |
Донная глубинная область Мирового океана |
Сложность очень малая |
|
Гидротермально-сиповый |
Температура высачиваний и флюида до сотен град.С. |
От малого до сильного |
Слабые и умеренные |
Сульфидные твердые постройки, илы |
Симбионты, бактериальные агрегации, маты, взвешенные в воде микроформы. |
Отсутствуют |
Седентарные и подвижные симбиотрофы |
Симбиотрофные и свободноживущие серобактерии |
Симбиотрофия, |
Рифтовые зоны, донные высачивания , островной характер распространения |
Высока сложность в зоне гидротермальных излияний, выражены концентрические структуры |
|
Лимнобиом |
От замерзающих до тропических водоемов. |
Замедленный |
Слабые, ветровые, сейши |
Водная среда как опора. Донные субстраты разной дисперсности и природы, включая высшие растения |
Взвешенные в воде микроформы. |
Взвешенные в воде микроформы, макрофиты. |
Взвешенные в воде, активно подвижные и седентарные мнзо- и макроформы |
Фотоавтотрофы, взвешенные в воде, прикрепленные микроформы, макрофиты |
Пастбищные и детритные трофические цепи |
Понижения рельефа суши, в которых накапливается вода поверхностного стока. |
Глубинная стратификация хорошо выражена. |
|
Реобиом |
От замерзающих до тропических водотоков. Широтная зональность выражена |
Очень высокий |
От слабого до сильного, всегда хорошо выражено. |
Донные субстраты различного качества, водная среда как опора. |
Микро- и макроформы на различных субстратах Взвешенные в воде микроформы. |
Седентарные микроформы. |
Седентарные, закапывающиеся микро- и макроформы |
Седентарные микроформы, макрофиты |
Пастбищные и детритные трофические цепи |
Понижения рельефа, по которому происходит поверхностный сток. |
Глубинная стратификация выражена слабо. |
Основными характеристиками среды в пелагическом океаническом биоме являются свойства водных масс океана: замедленный в целом водообмен (относительно водообмена поверхностных вод суши), существование фронтов и разномасштабных циркуляций, разделение на относительно малую по объему фотическую, изменчивую по температуре и большую по объему дисфотическую и достаточно стабильную по температуре холодную зоны. Однако, эти характеристики могут быть использованы для деления биома на меньшие категории, и не влияют на целостность всего биома (мегабиома). Характеристика жизненных форм океанического пелагического биома может быть образно охарактеризована так: его населяют организмы, которые "не знают" и "не подозревают" о существовании границ вода-берег, вода-дно. Переходя эту границу, например, переходя из стадии пелагической личинки к седентарной стадии взрослой особи, организмы попадают в другие экосистемы и в другой биом и представляют собой уже совершенно иные жизненные формы и экоморфы. Пелагический мегабиом океана может быть разделен на два макробиома – по характеру освещенности и, соответственно, с различными трофо-энергетическими характеристиками с автотрофно-гетеротрофными (фотическая зона) и гетеротирофными (афотическая) сообществами.
Прибрежный или циркумконтинентальный биом океана ограничен береговой линией и шельфовой зоной. Здесь тесное взаимодействие водной толщи, её поверхности, дна, граничной области море-суша создают чрезвычайно большое биотопическое разнообразие. Это является предпосылкой существования здесь многих жизненных форм гидробионтов – седентарных и подвижных, с разнообразными типами питания. Значительная часть экосистем этого биома автотрофно-гетеротрофные, высоко продуктивны. В прибрежном мегабиоме хорошо выражена широтная зональность, что позволяет выделять биогеографические прибрежные шельфовые области с различными условиями обитания и различающимся населением. Тесное взаимодействие с биомами суши проявляется в эвтуарных зонах.
Особенности генезиса, состава жизненных форм, метаболических процессов позволяют выделить биогермовый биом, хотя пространственно он частично совпадает с прибрежным. Биогермовым биомом представляет собой комплекс экосистем, в которых живыми организмами создаются структуры, сопоставимые с геоморфологическими, которые являются топической основой существования экосистем [45, 60]. Хотя в настоящее время этот биом представлен в основном коралловыми экосистемами, начиная с докембрийских времен не менее полутора десятков групп организмов (от цианобактерий до мшанок) участвовали в формировании биогермовых экосистем [45].
Батиально-абиссальный биом океана представляет собой совокупность донных экосистем за пределами шельфа. Топически, видимо, целесообразно выделить макробиомы – срединно-океанических хребтов, донных осадков океанического ложа, глубоководных желобов ультраабиссали. Основной особенностью этих экосистем является их полная энергетическая зависимость от процессов первичного продуцирования в фотической зоне океана. Именно поэтому, как отмечал Г.В.Богоров [10] минимальным изопланктам в поверхностной зоне соответствуют минимальные изобенты. Основными жизненными формами гидробионтов в этом биоме являются седентарные и подвижные с опорой на донный субстрат формы.
Деление как пелагического, так и донного океанического биомов по экологическим критериям совпадает в основных чертах с фаунистическим. В пелагическом биоме можно выделить мелководную (продуцирующую, 0-200 м), переходную – 200 - 1000 м– и глубоководную, потребляющую области [6].
Гидротермально-сиповый биом можно назвать также биомом восстановительных условий [8]. Основным абиотическим фактором выступает не столько высокая температура гидротермальных флюидов, сколько локальное преобладание восстановительных условий над окислительными. Особенностью этого биома как совокупности своеобразных экосистем является их островное расположение [14, 61]. Трофическая структура этих сообществ базируется на функционировании хемоавтотрофных организмов [15, 18] и значительную роль играют симбиотрофические связи.
Все экосистемы поверхностных вод суши при наиболее общем подходе могут быть разделены на две большие группы: лимнобиом – совокупность экосистем водоемов с замедленным водообменом и реобиом или совокупность экосистем текучих вод и. На уровне отдельных экосистем бывает сложно обособить экосистемы водоемов и водотоков.
Лимнобиом в масштабах континентов имеет мозаичную, островную природу. Водоемы с замедленным водообменом более или менее ограничены друг от друга, объединяются водотоками. Основной градиент условий проходит в них от литорали в глубину к профундальной части. В литоральной области преобладают седентарные формы, в том числе высшие водные растения.
Водотокам свойственна структура последовательных, связанных по их протяженности экосистем, обобщенный взгляд на которую представлен в концепции речного континуума [62]. Бывает практически невозможно выделение и разграничение отдельных малых экосистем, поэтому предлагается введение понятия реобиом [9]: "надо говорить о существовании особой надэкосистемной формы организации природных комплексов – континууме речных экосистем" (с.165). Очевидно, что такой подход может быть распространен не только на экосистему конкретной реки, но на всю речную сеть данного бассейна. В то же время, по основным признакам – общему характеру условий среды, ведущих жизненным форм гидробионтов, организации речного континуума все экосистемы водотоков в отличие от других гидроэкоситем могут быть объединены в единый мегареобиом гидросферы. Таким образом микро- и мезореобиомы представляют собой топо-хорологические единицы, а макрореобиом – как типологические, с позиций концептульного укрупнения понятий, так и хорологические – с позиций структуры биогидросферы.
Заключение
Биом и, в частности, гидробиом представляет собой категорию хорологическую. Совокупности сходных по своему строению экосистем занимают вполне определенное пространство. Биом выглядит как “ареал” сходных экосистем. Определенное сходство состава жизненных форм индицирует сходство комплекса условий существования организмов. Существует определенная структура биомов как хорологических единиц биосферы (Рис. 2).
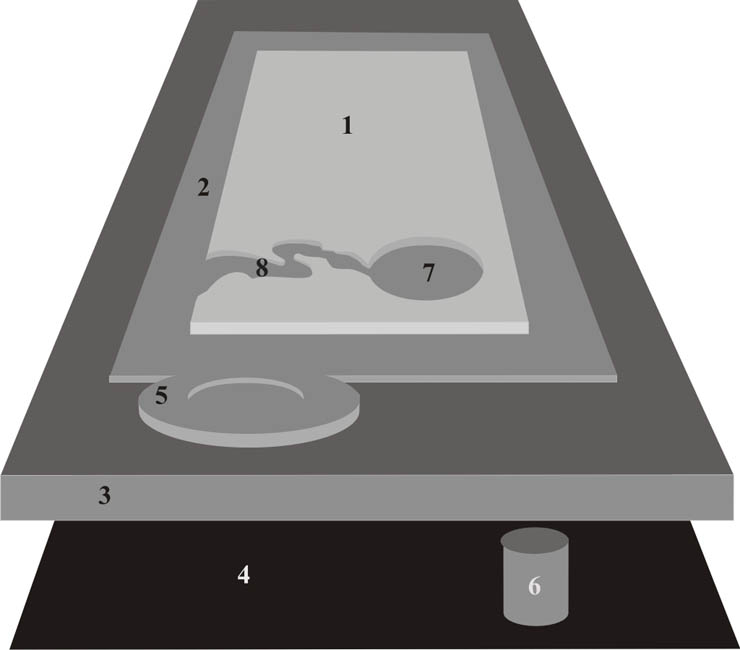
Рис. 2 Структура биосферы (биомы суши и гидросферы)
1- биомы суши, 2- прибрежный биом океана, 3- пелагический океанический биом, 4- батиально-абиссальный биом, 5- биогермовый биом, 6- гидротермально-сиповый биом, 7- лимнобиом, 8- реобиом.
Fig. 2. The structure of the Biosphere (biomes of land and hydrosphere)
1- land biomes, 2- coast ocean biome, 3- pelagic ocean biome, 4- bathial-abissal bottom biome, 5- biogerm biome, 6- vent-seep biome, 7- limnobiome, 8- rheobiome.
Биом также – категория экологическая и функциональная. Если идти от частного к общему, биом представляет собой систему сходных экосистем, надэкосистемное образование. При этом биомы неравноценны по роли биотического фактора. В этом аспекте может быть предложен, например, такой ряд по увеличению роли биоты: пустынный биом – пелагический биом океана – степной биом – прибрежный – гилея – биогермовый биом. С другой стороны (от общего – к частному), биосфера состоит из крупных функциональных блоков – биомов.
Кроме того, гидробиом – категория гидробиологическая. Если рассматривать гидробиологию как науку о закономерностях существования определенным образом структурированного живого вещества в гидросфере, то гидробиом выступает как наибольшая структурно-функциональная единица деления гидросферы с её населением.
Благодарности. Автор искренне признателен д.б.н. А.И.Кафанову, д.б.н. А.Ю.Звягинцеву, Институт биологии моря ДВНЦ, к.б.н. В.И. Юришинцу, к.б.н. Т.Н. Дьяченко, Институт гидробиологии НАНУ, за конструктивные советы при обсуждении материала работы, а также C. Rapin, EAWAG, Швейцария, за помощь в знакомстве с литературой.
Biomes in the hydrosphere. A.A. Protasov.
The concept of biome with reference to hydrosphere is considered, that has allowed to prove allocation of biome as structural element of Earth Biosphere as a whole. The hydrobiome as а eco-biogeographical, bioscherelogical and hydrobiological category is considered.Key words: biome, hydrosphere, Biosphere of the Earth, hydrobiology, biogeography of hydrosphere.
Біоми у гідросфері. О.О. Протасов. Розглянуто концепцію біому щодо гідросфери, що дозволило обгруноувати виділення біому як структурного елементу біосфери взагалом. Гідробіом розглядається як біосферологічна, еко-біогеографічна та гідробіологічна категорії.
Ключові слова: біом, гідросфера, біосфера Землі, гідробіологія, біогеографія гідросферы.