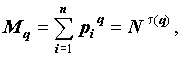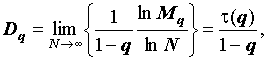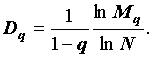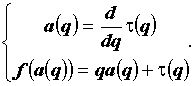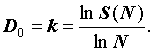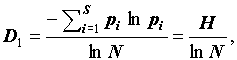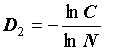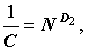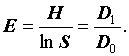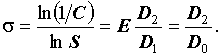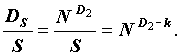© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic"
На главную страницу сайта
Сайт наш чаще посещай - будет выше урожай (Лозунг времен Н.С.Хрущева)
|
© 2004 Электронный журнал "Jahrbuch fur EcoAnalytic und EcoPatologic" |
|
Опубликовано: сб. докладов Всероссийской конференции "АКТУАЛЬНЫЕ ПРОБЛЕМЫ ВОДОХРАНИЛИЩ",2002, Борок, ИЭВВ РАН.
МУЛЬТИФРАКТАЛЬНЫЙ АНАЛИЗ ВИДОВОГО РАЗНООБРАЗИЯ ЗООПЛАНКТОЦЕНОЗОВ ЧЕБОКСАРСКОГО ВОДОХРАНИЛИЩА
Шурганова Г. В., Иудин Д. И., Гелашвили Д. Б., Якимов В. Н.
Нижегородский госуниверситет им. Н. И. Лобачевского, Нижний Новгород, Россия, ecology@unn.ac.ru
Ретроспективная гидробиологическая характеристика Чебоксарского водохранилища
С образованием Чебоксарского водохранилища завершилось создание крупнейшего в мире Волжского каскада водохранилищ. Заполнение водохранилища осуществлено в 1981 г. Общая длина водохранилища составляет 321 км, максимальная ширина – 16 км, средняя глубина – 4,2 м, площадь водного зеркала – 1080 км2 (Литвинов, 2000). Водохранилище отличается высоким коэффициентом водообмена (19,8), непостоянством уровенного режима и существенным загрязнением воды, наиболее сильно проявляющимся в его речной части. Формирование водных масс водохранилища происходит за счет разнородных водных потоков: левобережного, поступающего из Горьковского водохранилища, и правобережного, вносимого р. Окой (до 40% притока воды). Воды этих потоков имеют значительные различия гидрофизических и гидрохимических параметров: температуры, прозрачности, электропроводности и др. Правобережные воды отличаются более высоким содержанием биогенов, значительной минерализацией.
Речной участок водохранилища отличается тем, что лево- и правобережные водные потоки идут не смешиваясь на значительном протяжении. Достигая озерной части они перемешиваются и различия гидрофизических и гидрохимических параметров нивелируются.
Перечисленные особенности делают Чебоксарское водохранилище уникальным искусственным водоемом, не имеющим аналогов как в Волжском каскаде, так и в других каскадах водохранилищ Европы.
Гидробиологические исследования бактерио-, фито-, зоопланктона и зообентоса р. Волги на трассе строительства Чебоксарского водохранилища начались задолго до зарегулирования р. Волги у г. Чебоксары. Проводились регулярные комплексные мониторинговые исследования, продолжившиеся и в первое десятилетие существования водохранилища. Опубликовано достаточно большое количество работ, в которых приводятся сведения об изменениях видовой структуры, количественных характеристик гидробиоценозов на акватории Чебоксарского водохранилища (Шахматова и др., 1981; Шахматова, Разгулов, 1985; Тарасова и др., 1985; Тарасова, 1990; Охапкин, 1994 и др.).
Данная работа содержит результаты исследования зоопланктона р. Волги на трассе строительства Чебоксарского водохранилища в “доводохранилищный” период, а также зоопланктона Чебоксарского водохранилища – с момента его образования (1981 г.) по 2002 г. В первые двенадцать лет существования водохранилища проводились регулярные мониторинговые исследования в составе комплексных экспедиций. Пробы зоопланктона отбирались в разные сезоны каждого года (весна, лето, осень) в лево- и правобережье водоема на 18 стационарных станциях. В последующие годы проводились отдельные съемки по всей акватории водохранилища. Для сбора материала использовалась количественная сеть Джеди (диаметр входного отверстия 18 см, капроновое сито №64). Отбор осуществлялся путем тотальных ловов от дна до поверхности. Обработка материалов проводилась общепринятым в гидробиологии счетно-весовым методом (Киселев, 1969; Методические рекомендации …, 1984). Идентификацию видов проводили с использованием определителей зоопланктона (Кутикова, 1970; Мануйлова, 1964; Определитель …, 1995; Смирнов, 1971; Рылов, 1948).
В результате многолетних исследований выявлен видовой состав, сезонная динамика численности и биомассы, соотношение основных групп, оценено видовое разнообразие зоопланктоценозов Чебоксарского водохранилища. Проведен анализ динамики и пространственного распределения показателей видовой структуры с помощью многофакторного регрессионного анализа. Выявлены тенденции пространственного распределения основных зоопланктоценозов водохранилища на начальных этапах и в ходе экзогенной сукцессии. Высокий уровень водообмена, непостоянство гидрологического режима, а также загрязнение воды вызывают активно идущие процессы антропогенной трансформации зоопланктоценозов на протяжении всего периода существования водохранилища. Анализ всех исследуемых показателей видовой структуры зоопланктоценозов водохранилища за более чем двадцатилетний период его существования свидетельствует о сложности протекающих в водохранилище процессов, существовании на его акватории отдельных зоопланктоценозов, состояние которых может быть охарактеризовано как нестационарное (Шурганова, 1984, 1986, 1987, 1989, 1996, 2002; Шурганова, Кузнецова, 1984, 1985, 1987; Шурганова, Черников, 1989, 1990; Шурганова и др., 1997; Шурганова, Махин, 1998; Кузнецова и др., 1991 и др.).
Для получения более полного, системного представления о зоопланктоценозах Чебоксарского водохранилища при проведении детального анализа их видовой структуры за более чем двадцатилетний период его существования нами был использован мультифрактальный анализ. Предлагаемый метод, широко применяемый в физических и математических исследованиях для описания сложных систем и недавно предложенный для анализа структуры биотических сообществ (Иудин, Гелашвили, 2002; Гелашвили, Розенберг, 2002), впервые был применен в гидробиологических исследованиях для планктонных сообществ крупных равнинных водохранилищ.
Фракталы и мультифракталы в гидробиологии
Самоподобие является весьма общим свойством природных систем: бассейны крупных рек, ветвящиеся каналы молниевого разряда, пространственная структура колоний микроорганизмов, распределение звездного вещества в космосе – все эти и многие другие сильно неравновесные системы, связанные, так или иначе, с переносом интенсивных потоков вещества, заряда, энергии, – обладают удивительной структурной универсальностью. Часто в этой связи говорят о фрактальности природных объектов (Mandelbrot, 1972; Bunde, Havlin, 1994).
В настоящем исследовании мы обратимся к анализу самоподобия видовой структуры биотических сообществ. На фоне физических проявлений фрактальности биоценозов (Азовский, Чертопруд, 1998; Медвинский и др., 2002) проблема самоподобия видовой структуры весьма специфична. Дело в том, что, если физические проявления самоподобия очевидным образом соотносятся с самоподобием соответствующих геометрических носителей, – ручейков, каналов, пространственно-временных распределений и т.д., то структурные особенности внутренних энергетических каналов и иерархия связей биотических сообществ остаются скрытыми от глаз: они не имеют непосредственных геометрических образов инвариантных относительно преобразований масштаба. Тем не менее, они демонстрируют самоподобие при изменении численности сообщества. Одним из первых этот фундаментальный факт отметил Маргалеф (Маргалеф, 1992). Именно он указал на степенную зависимость числа видов S от численности сообщества N
|
|
(1) |
где 0 ≤ k ≤ 1 – было предложено рассматривать как индекс разнообразия. Выражение (1) показывает, что видовая структура сообщества инвариантна относительно преобразования его численности. Величину k можно трактовать как соответствующую фрактальную размерность: число элементов видовой структуры – число видов сообщества – меняется по степенному закону с показателем k при увеличении размеров системы, т.е. с ростом численности сообщества.
Однако, как и любая фрактальная размерность, индекс k не дает исчерпывающего количественного представления о видовой структуре биотического сообщества: выражение (1) констатирует лишь сам факт наличия вида в выборке, но не содержит, например, сведений о распределении видов по численности или о степени их доминирования. Наиболее общее описание внутреннего устройства самоподобных объектов позволяет дать теория мультифракталов, характеризуемых бесконечной иерархией размерностей (Федер, 1991).
Рассмотрим отличия монофрактального и мультифрактального подходов. Главной характеристикой фрактального множества является его размерность. Однако стандартная процедура определения фрактальной размерности не позволяет обнаруживать различия между однородными и неоднородными (мультифрактальными) объектами. Она заключается в том, что исходный объект покрывается областями (ячейками) меньшего масштаба l. Затем подсчитывается число ячеек n(l), в которых содержится хотя бы одна точка изучаемого объекта, и уже эти значения используются в дальнейшем анализе. Поэтому данная процедура нечувствительна к неоднородности распределения точек между ячейками.
Таким образом, использование обычной фрактальной размерности не позволяет отличить однородные объекты от неоднородных. Для этого вводятся новые характеристики. При этом процедура определения размерности несколько усложняется: для каждой заполненной ячейки подсчитывается число содержащихся в ней точек 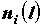 , которое затем преобразуется в долю
, которое затем преобразуется в долю 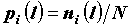 , где N – общее число точек. Далее используется полученный набор {pi}.
, где N – общее число точек. Далее используется полученный набор {pi}.
Введем моменты распределения точек по ячейкам:
|
|
(2) |
где -∞ ≤ q ≤ ∞ – называется порядком момента, а показатель 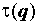 характеризует скорость изменения соответствующего момента при увеличении размера выборки.
характеризует скорость изменения соответствующего момента при увеличении размера выборки.
Обобщенной размерностью Dq (обобщенной размерностью Реньи) распределения называется функция порядка момента q, вводимая определением (Федер, 1991):
|
|
(3) |
На практике величины (3) можно оценить используя несколько различающихся значений N по более простой формуле:
|
|
(4) |
В общем случае можно получить бесконечное число обобщенных размерностей Dq, поэтому неоднородный фрактальный объект называют мультифракталом. Для однородных фракталов 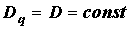 , поэтому их часто также называют монофракталами.
, поэтому их часто также называют монофракталами.
Свойства мультифрактала полностью описываются характерным для него набором обобщенных фрактальных размерностей Dq, который, в свою очередь, может быть представлен в виде некоторой нелинейной функции 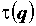 . Однако величины Dq не являются, строго говоря, фрактальными размерностями в обычном понимании. Поэтому наряду с ними используется так называемая функция мультифрактального спектра f(a). Ее получают путем преобразования Лежандра функции
. Однако величины Dq не являются, строго говоря, фрактальными размерностями в обычном понимании. Поэтому наряду с ними используется так называемая функция мультифрактального спектра f(a). Ее получают путем преобразования Лежандра функции 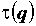 :
:
|
|
(5) |
Преимущество функции мультифрактального спектра состоит в том, что ее значения представляют собой размерности неких однородных фрактальных подмножеств из исходного множества, которые дают доминирующий вклад в моменты распределения при заданных q. Кроме того, график функции f(a) компактен и не уходит на бесконечность.
Обратимся теперь к существующим аналогиям между фрактальным анализом геометрических объектов и анализом структуры биологических сообществ. В обоих случаях занимаемое объектом пространство разбивается на ячейки. В случае биологического сообщества такое разбиение происходит неявно, так как для элементов сообщества все пространство возможностей изначально разбито на таксономические категории (чаще всего в качестве таких ячеек выступают виды). Подсчет числа составляющих сообщество видов ( то есть определение видового богатства сообщества) аналогичен подсчету занятых ячеек при определении размерности однородного фрактала. Таким образом, анализ видового богатства сообщества можно сопоставить с монофрактальным подходом к анализу неоднородных объектов.
Как биологическое сообщество, так и геометрический мультифрактал обладают гетерогенностью (в сообществах, например, выделяются доминантные и редкие виды), поэтому при простом подсчете занятых ячеек теряется важная информация. Чтобы этого избежать, биологи переходят к анализу видового разнообразия сообществ, что аналогично переходу к мультифрактальному анализу. В обоих случаях это сводится к определению содержимого каждой из ячеек в отдельности. Итогом является описанный выше набор долей {pi}. Для сообщества pi – это доля особей i-го вида. Далее, исходя из этого набора, можно вычислить различные индексы видового разнообразия (Шеннона, Симпсона и др.).
Покажем взаимосвязь различных индексов разнообразия с использованием мультифрактального анализа. Рассмотрим выражения для некоторых обобщенных размерностей Dq.
Для q = 0 результат очевиден:
|
|
(6) |
Можно показать, что для q = 1:
|
|
(7) |
где 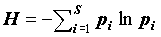 есть ни что иное как информационный индекс видового разнообразия Шеннона-Уивера (Shannon, Weaver, 1949). Для q = 2:
есть ни что иное как информационный индекс видового разнообразия Шеннона-Уивера (Shannon, Weaver, 1949). Для q = 2:
|
|
(8) |
или
|
|
(9) |
где 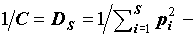 известный в экологической литературе индекс видового разнообразия Симпсона (Simpson, 1949; Алексеев и др., 1992).
известный в экологической литературе индекс видового разнообразия Симпсона (Simpson, 1949; Алексеев и др., 1992).
Обобщенные размерности практически не зависят от численности сообщества и являются своеобразными структурными инвариантами однотипных систем отличающихся друг от друга размерами.
В отличие от них, часто используемые индексы Симпсона (C, DS) и Шеннона (H) зависят и от численности и от числа видов: 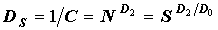 и
и 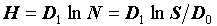 . Следовательно, сами по себе, они не могут охарактеризовать разнообразия всего сообщества в целом и требуют дополнительной нормировки. Так, например, разделив индекс Шеннона на логарифм числа видов, получим индекс эквитабильности Пиелу (Pielou, 1966), который уже не зависит от числа видов и инвариантен относительно роста размеров системы:
. Следовательно, сами по себе, они не могут охарактеризовать разнообразия всего сообщества в целом и требуют дополнительной нормировки. Так, например, разделив индекс Шеннона на логарифм числа видов, получим индекс эквитабильности Пиелу (Pielou, 1966), который уже не зависит от числа видов и инвариантен относительно роста размеров системы:
|
|
(10) |
В случае с индексом видового разнообразия Симпсона DS, нормировка его логарифма на логарифм числа видов дает новый индекс разнообразия σ, который также является инвариантом:
|
|
(11) |
Обобщенную размерность D2 называют в теории фракталов корреляционной размерностью (Goltz, 1998), поэтому индекс разнообразия σ можно назвать корреляционным инвариантом. Заметим, что каноническая форма индекса эквитабильности Симпсона 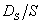 инвариантом не является:
инвариантом не является:
|
|
(12) |
Непосредственной проверкой легко убедиться в том, что каждый из всего бесконечного множества инвариантов вида 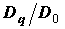 , представляет собой индекс эквитабильности, и меняется в пределах от нуля (для хемостата) до единицы (в случае равнопредставленности видов). Заметим, что индекс Маргалефа k (впрочем, как и все обобщенные размерности Dq) достигает единицы только при S = N.
, представляет собой индекс эквитабильности, и меняется в пределах от нуля (для хемостата) до единицы (в случае равнопредставленности видов). Заметим, что индекс Маргалефа k (впрочем, как и все обобщенные размерности Dq) достигает единицы только при S = N.
Итак, любое биологическое сообщество можно рассматривать как мультифрактальный объект и характеризовать его спектром размерностей. Дадим биологическую интерпретацию различным частям мультифрактального спектра.
Следует сразу отметить, что при таком подходе сообщество рассматривается как множество, состоящее из отдельных фрактальных подмножеств, которые можно интерпретировать как совокупности особей, относящихся к видам со сходной представленностью. Так, например, можно выделить подмножество доминирующих видов, подмножество субдоминантов и т.д. Для таких подмножеств можно вычислить фрактальную размерность, которая и будет характеризовать видовое разнообразие. Именно такой смысл имеет ордината точек на графике мультифрактального спектра. Абсцисса же точек характеризует представленность (долю) видов того или иного подмножества. Ее также можно интерпретировать как скорость убывания доли вида (типичного представителя данного подмножества) при росте объема выборки. Наличие на графике спектра точек, лежащих по оси “а” близко к нулю, означает присутствие в сообществе сильных доминантов (причем, чем меньше абсцисса точек, тем сильнее доминирование); наличие же точек, лежащих близко к единице, означает присутствие в сообществе редких видов. Таким образом, ширину спектра можно интерпретировать как показатель выровненности видов в сообществе: чем шире спектр, тем меньше выровненность, и наоборот. Крайним вариантом выровненности является равнопредставленность видов; при этом весь спектр схлопывается в единственную точку, ордината которой соответствует монофрактальной размерности сообщества.
Из сказанного следует, что точки кривой мультифрактального спектра описывают разнообразие в отдельных группах видов. В то же время, на этой кривой находятся точки, которым можно поставить в соответствие известные индексы разнообразия, характеризующие сообщество в целом. Однако в этом нет непреодолимого противоречия. Дело в том, что различные индексы разнообразия придают разное значение видам с отличающейся представленностью. Так, например, широко известно, что основной вклад в индекс разнообразия Симпсона вносят широко представленные виды. Поэтому различные индексы разнообразия действительно в большей степени характеризуют отдельные группы видов с определенной представленностью. Для получения же полной информации о структуре сообщества необходимо рассмотрение всего спектра индексов. Такую возможность и предоставляет мультифрактальный анализ.
Мультифрактальная структура зоопланктоценозов Чебоксарского водохранилища
Как показано в предшествующих работах (Шурганова, 1986, 1996, 2002 и др.), разнородным водным массам Чебоксарского водохранилища соответствуют отдельные зоопланктоценозы, меняющиеся в ходе экзогенной сукцессии. Для оценки сложности структуры этих ценозов был применен мультифрактальный анализ.
Было выделено четыре зоопланктоценоза, расположенных в лево- и правобережье речной части, а также в лево- и правобережье озерной части водохранилища. Для каждого из этих ценозов были суммированы данные проб соответствующих станций и построены мультифрактальные спектры.
Вид полученных спектров значительно различается для сообществ зоопланктона на разных участках водохранилища в один период времени. Значительно отличаются мультифрактальные спектры и на одном и том же участке водохранилища в разные моменты времени.
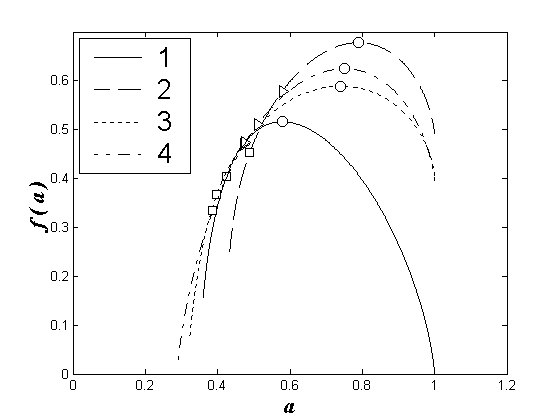
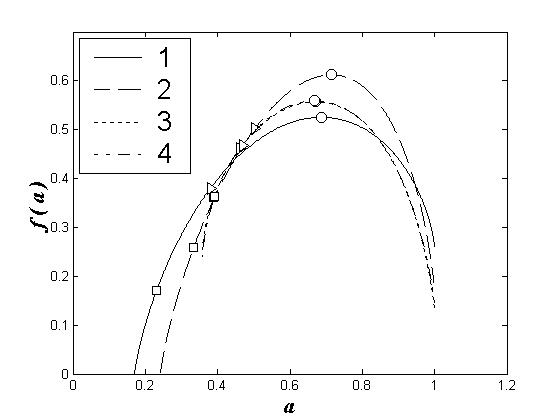
Покажем это на примерах (см. рис. 1).
|
|
|
|
|
|
Рис. 1. Мультифрактальные спектры видовой структуры зоопланктоценозов Чебоксарского водохранилища: 1 – левобережный речной участок, 2 – правобережный речной участок, 3 – левобережный озерный участок, 4 – правобережный озерный участок.
o
– соответствуют обобщенным размерностям D2, ê
– обобщенным размерностям D1, ¡
– обобщенным размерностям D2. Обозначения размерностей сохраняются для рис. 2-3.
В доводохранилищный период заметны значительные различия спектров, характеризующих лево- и правобережные сообщества зоопланктона (рис.1а), что объясняется значительной разнородностью водных масс, формирующих исследуемый водоток, и различием расположенных в этих водах сообществ зоопланктона. Правобережный водный поток, несущий окские воды, содержит реофильный окский зоопланктон. Левобережный водный поток, поступающий из приплотинного плеса Горьковского водохранилища, содержит лимнофильный комплекс трансформированного планктона.
В период формирования водохранилища (1981 – первый год его существования) в образующейся озерной части наблюдалось значительное перемешивание водных масс, что обусловило относительную однородность зоопланктоценозов. Вследствие этого мультифрактальные спектры для сообществ лево- и правобережья озерной части водохранилища полностью совпадают. В то же время, мультифрактальные спектры для сообществ зоопланктона лево- и правобережья речной части водохранилища в этот период сохраняют значительные различия (рис. 1б).
В период относительной стабилизации зоопланктона водохранилища (после 1985 г.) различия мультифрактальных спектров для сообществ лево- и правобережья речной части водоема сохраняются, однако их степень уменьшается по сравнению с доводохранилищным периодом. Сходство спектров для зоопланктоценозов озерной части водохранилища становится менее отчетливым по сравнению с таковым в период формирования водохранилища (рис. 1в).
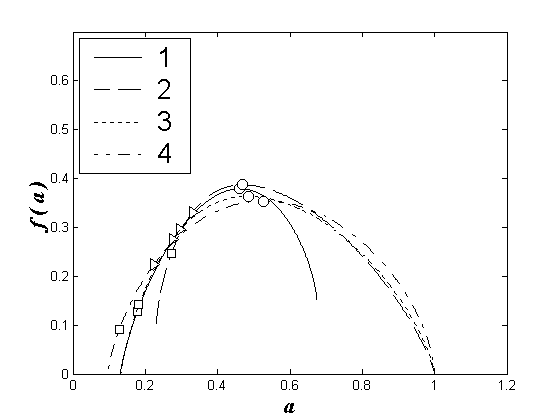
Аналогичная картина наблюдается и в 2002 г. для мультифрактальных спектров, построенных для лево- и правобережных станций у городов Нижний Новгород и Чебоксары (рис. 2).
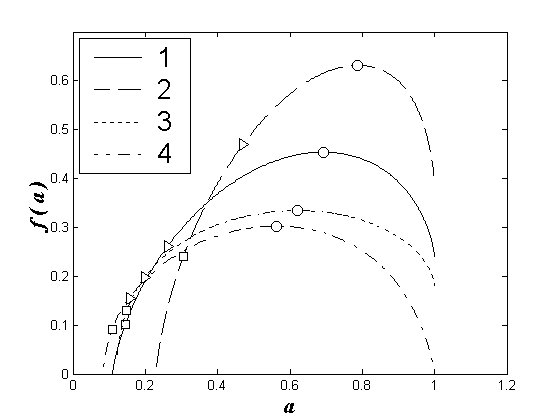
Рис. 2. Мультифрактальные спектры видовой структуры зоопланктоценозов Чебоксарского водохранилища для июля 2002 года. 1 – Нижний Новгород, левобережье, 2 – Нижний Новгород, правобережье, 3 – Чебоксары, левобережье, 4 – Чебоксары, правобережье
Проведем анализ мультифрактальных спектров одного из участков водохранилища (правобережного речного) в весенние периоды ряда лет наблюдения. Зоопланктоценоз этого участка в последний год перед созданием Чебоксарского водохранилища (1979) в весенний период характеризовался присутствием двух доминантов – коловраток Keratella quadrata (Muller, 1786) и Brachionus calyciflorus Pallas, 1766. В последующие годы с возникновением водохранилища на этом участке (правобережье средней речной части) в весенний период единственным доминантом стал Brachionus calyciflorus, вследствие чего произошло усиление степени доминирования.
Эти изменения отражены на рис. 3. Так, кривая мультифрактального спектра 1979 г. не достигает оси абсцисс, что свидетельствует о наличии более чем одного доминирующего вида (в данном случае – двух). После образования водохранилища (1981, 1983, 1987 гг.) произошло смещение кривых по оси “а” влево, к нулю, что означает усиление степени доминирования в сообществе. Указанные кривые стали также стремиться к оси абсцисс, что означает наличие лишь одного доминирующего вида в зоопланктоценозе.
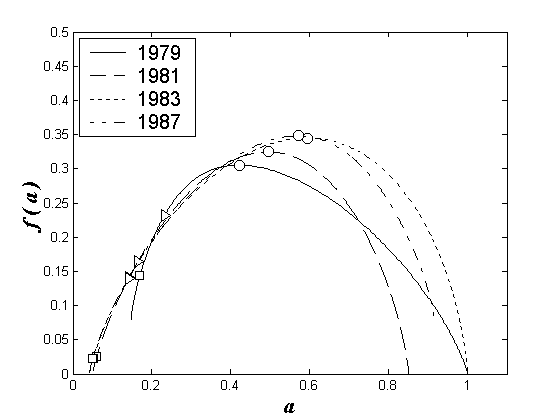
Рис. 3. Мультифрактальные спектры видовой структуры зоопланктоценозов правобережья речной части Чебоксарского водохранилища для ряда лет
Мы видим, таким образом, что видовая структура биотических сообществ может быть охарактеризована с помощью мультифрактального формализма. Важно отметить, что мультифрактальный спектр распределения особей по видам объединяет в себе фрактальные размерности распределения с индексами сингулярности отдельных видов, что существенно расширяет возможности традиционного анализа видового разнообразия. Дальнейшее развитие мультифрактального подхода, в частности, выявление связи между параметрами мультифрактального спектра и биотическими и абиотическими факторами, является одной из актуальных задач экологии.
Авторы выражают глубокую признательность А.П. Левичу за полезное обсуждение работы и конструктивные замечания, учтенные авторами при подготовке публикации.
Работа выполнена при поддержке Российского фонда фундаментальных исследований, грант 01-02-17403.
Литература
Азовский А.Н., Чертопруд М.В. Масштабно-ориентированный подход к анализу структуры сообществ // Журн. общ. биол. 1998. Т. 59. №52. С. 117-136.
Алексеев В.В., Крышев И.И., Сазыкина Т.Г. Физическое и математическое моделирование экосистем. – СПб.: Гидрометеоиздат, 1992. – 368 с.
Гелашвили Д.Б., Розенберг Г.С. Фрактальная организация экосистем разной масштабности // Вопросы практической экологии. Материалы научной конференции. Пенза, 2002. С. 42-43.
Иудин Д.И., Гелашвили Д.Б. Применение мультифрактального анализа структуры биотических сообществ в экологическом мониторинге // Проблемы регионального экологического мониторинга. Материалы научной конференции. Нижний Новгород, 2002. С. 49-52.
Киселев И.А. Планктон морей и континентальных водоемов. Л., 1969. Т. 1.
Кузнецова М.А., Шурганова Г.В., Черников А.А. Анализ процессов трансформации зоопланктоценозов при зарегулировании стока с помощью показателей видового разнообразия // Экология. 1991. №4. С. 68-72.
Кутикова Л.А. Коловратки фауны СССР // Тр. Зоол. ин-та АН СССР. 1970. Вып. 104.
Литвинов А.С. Энерго- и массообмен в водохранилищах Волжского каскада. Ярославль, 2000.
Мануйлова Е.Ф. Ветвистоусые рачки (Cladocera) фауны СССР. М.; Л., 1964.
Маргалеф Р. Облик биосферы. – М.: Наука, 1992. – 254 с.
Медвинский А.Б., Петровский С.В., Тихонова И.А, Тихонов Д.А. Формирование пространственно-временных структур, фракталы и хаос в концептуальных экологических моделях на примере динамики взаимодействия популяций планктона и рыбы // УФН. 2002. Т. 172. №1. С. 31-66.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах // Зоопланктон и его продукция. Л., 1984.
Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 2. СПб., 1995.
Охапкин А.Г. Фитопланктон Чебоксарского водохранилища. Тольятти, 1994. 173 с.
Рылов В.М. Cyclopoida пресных вод. Фауна СССР. Ракообразные Т. 3., вып. 3. М.; Л., 1948.
Смирнов Н.Н. Chydoridae фауны мира. Фауна СССР. Ракообразные Т. 1., вып. 2. Л., 1971.
Тарасова Т.Н. Динамика численности и биомассы бактериопланктона Чебоксарского водохранилища по данным 1981-1985 гг. // Наземные и водные экосистемы: Межвуз. Сб. Горький, 1990. С. 71-76.
Тарасова Т.Н., Охапкин А.Г., Тухсонова Н.Г. Особенности формирования бактерио- фитопланктона и гидрохимического режима Чебоксарского водохранилища в первые годы наполнения // Формирование кормовой базы и ихтиофауны во вновь созданных водохранилищах Волжско-Камского каскада. Л., 1985. С. 14-22.
Федер Е. Фракталы. – М.: Мир, 1991. – 214 с.
Шахматова Р.А., Разгулов Ю.Н. Донная фауна Чебоксарского водохранилища в начальный период его существования // Формирование кормовой базы и ихтиофауны во вновь созданных водохранилищах Волжско-Камского каскада. Л., 1985. С. 27-34.
Шахматова Р.А., Тухсонова Р.Г., Охапкин А.Г., Шурганова Г.В., Разгулов Ю.Н. Комплексное изучение Волги на участке строительства нового водохранилища // IV съезд ВГБО. Тез докл. Ч. 4. Киев, 1981. С. 170-171.
Шурганова Г.В. Зоопланктон реки Волги на участке Городец – Чебоксары до образования нового водохранилища // Биологическая продуктивность и качество воды Волги и ее водохранилищ. М., 1984. С. 181-183.
Шурганова Г.В. Структурные изменения в планктонном ценозе водохранилища как показатель антропогенного воздействия // Наземные и водные экосистемы: Межвуз. сб. Горький, 1986. С. 26-32.
Шурганова Г.В. Динамика видовой структуры зоопланктонного сообщества в процессе его формирования (на примере Чебоксарского водохранилища). Автореф. дис. … канд. биол. наук. М., 1987.
Шурганова Г.В. Структурная характеристика основных зоопланктоценозов Чебоксарского водохранилища при промежуточном режиме его заполнения // Наземные и водные экосистемы: Межвуз. сб. Горький, 1989. С. 4-10.
Шурганова Г.В. Многолетняя динамика видовой структуры зоопланктоценозов водохранилища в условиях антропогенного пресса // Материалы VII съезда гидробиологического общества РАН. Казань, 1996. Т. 1. С. 223-224.
Шурганова Г.В. Изменения видовой структуры зоопланктоценозов Чебоксарского водохранилища в многолетнем аспекте // Актуальные проблемы водохранилищ. Тез. докл. Всероссийской конф. с участием специалистов из стран ближнего и дальнего зарубежья. 29 октября – 3 ноября 2002 г., Борок, Россия: Тез. докл. – Ярославль, 2002. С. 329-330.
Шурганова Г.В., Кузнецова М.А. Структура зоопланктона реки Волги на участке образования Чебоксарского водохранилища // Наземные и водные экосистемы: Межвуз. сб. Горький, 1984. С. 54-60.
Шурганова Г.В., Кузнецова М.А. Зоопланктон Чебоксарского водохранилища в первые годы его существования // Формирование кормовой базы и ихтиофауны во вновь созданных водохранилищах Волжско-Камского каскада. Л., 1985. С. 23-26.
Шурганова Г.В., Кузнецова М.А. Динамика основных зоопланктоценозов Чебоксарского водохранилища в первые годы его существования // Наземные и водные экосистемы: Межвуз. сб. Горький, 1987. С. 4-7.
Шурганова Г.В., Махин П.В. Динамика видовой структуры зоопланктоценозов речного участка Чебоксарского водохранилища в многолетнем аспекте // Экологические проблемы бассейнов крупных рек – 2. Тез. докл. междунар. конфер. Тольятти, 1998. С. 266-267.
Шурганова Г.В., Черников А.А. Статистическая оценка факторов, определяющих динамику биомассы зоопланктона Чебоксарского водохранилища в промежуточный период заполнения // Моделирование динамики популяций: Межвуз. сб. Горький, 1989. С. 7-11.
Шурганова Г.В., Черников А.А. Исследование динамики биомассы планктонных ракообразных Чебоксарского водохранилища с помощью статистических методов // Моделирование динамики популяций: Межвуз. сб. Н.Новгород, 1990. С. 44-48.
Шурганова Г.В., Черников А.А., Захаров Ю.К. Исследования динамики биомассы ракообразных Чебоксарского водохранилища с помощью статистических методов // Проблемы биологического разнообразия водных организмов поволжья. Зоопланктон, зообентос. Материалы научн. конфер. Тольятти, 1997. С. 72-76.
Bunde A., Havlin S. (Eds.) Fractals in Science, Springer-Verlag, Berlin, 1994.
Goltz C. Fractal and Chaotic Properties of Earthquakes, Springer, 1998.
Mandelbrot B.B. Possible refinement of the lognormal hypothesis concerning the distribution of energy dissipation in intermittent turbulence. – In: Statistical Models and Turbulence, eds. M. Rosenblatt and C. Van Atta, Lecture Notes in Physics 12, Springer, New York, pp. 333-351, 1972.
Pielou E.C. The measurement of diversity in different types of biological collections // J. Theor. Biol. 1966. V. 13. P. 131-144.
Shannon C., Weaver W. The Mathematical Theory of Communication. Urbara, Univ. of Illinois Press, 1949. 117 p.
Simpson E.H. Measurement of diversity // Nature. 1949. V. 163. P. 688.
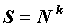 ,
,