| Дальше
|
Назад |
Начало |
Конец |
Список
|
4.3. Оценка качества экосистемы по индексам видового разнообразия
Гипотезы, используемые для оценки биоразнообразия
Популяционная и видовая плотность сообществ гидробионтов может меняться во времени в зависимости от изменений факторов среды. Это может происходить как в ходе эволюционных процессов, протекающих в течение длительного времени, так и при антропогенном эвтрофировании и загрязнении водоемов, когда процессы изменения структуры экосистем протекают с большими скоростями. Сокращается число видов, возрастает доминирование отдельных видов, для которых характерны более короткие жизненные циклы, проявляется раннее наступление половозрелости, увеличение биомассы и продукции. Ю. Одум [1986] отмечает, что при эвтрофировании и загрязнении водоемов преимущества получают эврибионтные виды с r-стратегией; в то время как в олиготрофных условиях незагрязненных водоемов, где разнообразие высокое и доминирование менее резко выражено, напротив, большее значение приобретают стенобионтные виды с длительными циклами развития и K-стратегией.
Видовое разнообразие слагается из двух компонентов [Одум, 1986]:
Вряд ли следует слишком буквально понимать термин “имеющиеся виды” без соответствующего уточнения. “Точечные разнообразия, т.е. разнообразия проб, – писал Р. Маргалеф [1992], – взятых в точке, почти всегда бессмысленны… Так как организация, выраженная через разнообразие, развертывается в пространстве, измерения не имеют большого смысла, если они не представлены в виде спектра: разнообразие как функция пространства” (можно добавить, "и времени"). Пусть в каждой точке территории ("столбике" по Р. Маргалефу) для каждого вида определена вероятность его возможного появления (такая вероятность может быть существенно мала, но никогда не превращается в нуль). Тогда под видовым богатством можно понимать количество видов, вероятность появления которых в окрестностях r данной точки больше a priori заданного числа e (e – параметр отбора). Естественно, видовое богатство растет с увеличением r (см. рис. 4.1), однако этот рост асимптотически стремится к некоторому порогу насыщения, характерному для каждого региона.
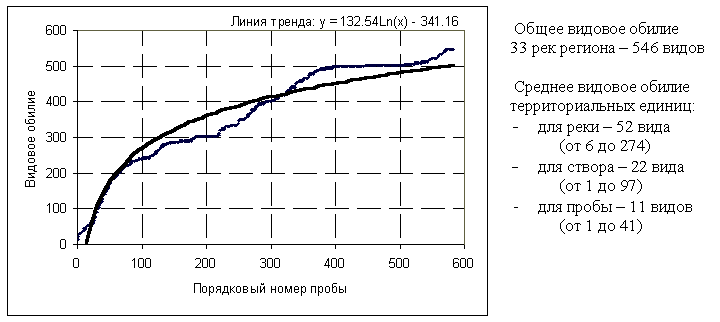
Рис. 4.1. "Открываемость" видов зообентоса по мере развития экспедиционных исследований малых рек Самарской области в 1991-2001 гг. (номер пробы выстроен в хронологическом порядке)
Таким образом, один из главных компонентов биоразнообразия – видовое богатство или плотность видов – это просто общее число видов, которое в сравнительных целях иногда выражается как отношение числа видов к площади или числа видов к числу особей. Так, например, Р.Маргалеф [Margalef, 1958БМ], исходя из того, что число видов пропорционально логарифму изученной площади [Gleason, 1922М], и считая, что общее число особей пропорционально площади, предложил в качестве меры биоразнообразия индекс видового богатства Маргалефа:
d = (s - 1) / ln N , (4.3)
(4.3)
где
s – число видов, N – число особей.Е.Ф. Менхиникк [Menhinick, 1964БМ] рассчитывал видовое богатство полевых насекомых по несколько другой формуле, используя в знаменателе функцию квадратного корня (индекс Менхиникка
):d
М = (s - 1) / (N)1/2 .Виды, входящие в состав биоценоза, очень сильно различаются по своей значимости. Традиционно принято выделение следующей иерархии видов: руководящие (или "доминантные") виды; за ними следует группа "субдоминантов"; остальные же виды считаются второстепенными, среди которых отмечают случайные или редкие. Значение отдельных видов должно определяться тем, какую роль играют они в функционировании экосистемы или в продукционном процессе. Но при исследованиях водных сообществ установить истинную функциональную роль видов нелегко, если об их значении судить только по обилию, т.е. численности и биомассе.
При этом для анализа биоразнообразия и степени доминантности в разных ситуациях используют два традиционных подхода:
Наилучший способ представить оба компонента разнообразия (т.е. богатство и выравненность) – построить график, где по оси ординат в логарифмическом масштабе отложено число особей, биомасса или иной рассматриваемый показатель, а по оси х
– ранжированная последовательность видов от наиболее до наименее обильного (см. рис. 4.2).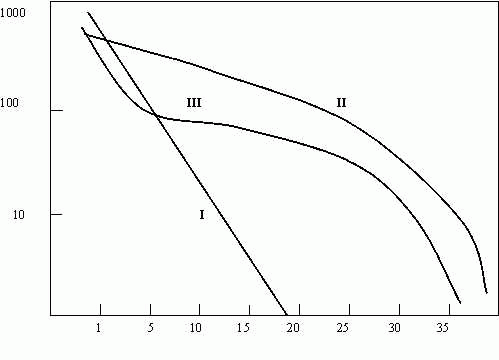
Рис.4.2. Форма кривых доминирования-разнообразия
Полученная в 1965 г. кривая названа Р. Уиттекером [1980] “кривой доминирования - разнообразия”, а Э. Пианкой [Pianka, 1978Б] – “кривой значимости видов”. Чем выше кривая и чем более она "уплощена", тем больше при данном числе видов их разнообразие. Считается, по форме этой кривой можно не только оценить видовую выравненность популяций, но и функциональную роль отдельных видов, их статус и биотический потенциал, т.е. все то, что достаточно расплывчато именуется “экологической нишей”. Так, для кривых на рис. 4.2 предлагается следующее толкование (S – число видов в пробе):
n(k) = n1 * Сk(k-1) ,
где
n(k) – значимость вида k в ряду от k = 1 (наиболее значимый вид) до k = S,С
k(k-1) = n(i) / n(i - 1);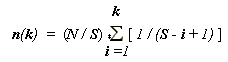 ,
,
где
N = å n(i)S
= å Sr = S0 * (p / a )0.5 ; Sr = S0 * exp[-(aR)2] ,где
Sr – число видов в октаве, удаленной на R октав от модального интервала, содержащего S0 видов; а @ 0,2 – постоянная, связанная со стандартным отклонением данного распределения.Гипотеза Ф. Престона порождает большое семейство S-образных кривых и соответствует промежуточной ситуации между I и II, наиболее широко распространенной в природе.
Индексы доминирования
Рассмотрим некоторые используемые выражения для индексов, позволяющих выделить виды-доминанты. Например, для природных биоценозов принято использовать индекс доминирования И. Балога [1958, цит. по: Руководство по гидробиологическому.., 1992]:
Di = Ni / Ns,
где
Ni – число особей i-го вида, Ns – общее число особей в биоценозе. К сожалению, этот идеальный по своей простоте индекс не отражает самого смысла доминирования, поскольку может принимать, например, значение 0.5 как при истинном доминировании, когда при нескольких сотнях видов один вид выражен половиной численности, так и в случае двух особей двух видов.Другая формула индекса доминирования (или доминантности) предложена А. Ковнацки [Kowna
cki, 1971М] на основе "коэффициента обилия" В.Ф. Палия [1961] (индекс доминирования Палия - Ковнацки):Di
= 100 × pi × Ni / Ns , (4.4)
где
pi – встречаемость; pi = mi / Mi , mi – число проб, в которых был найден вид i, M – общее число проб, Ni – число особей i-го вида, Ns – общее число особей в биоценозе. Для характеристики видового комплекса предлагается выделять доминанты в пределах 10 < Di < 100, субдоминанты – в пределах 1 < Di < 10, субдоминанты первого порядка – в пределах 0.1 < Di < 1 и второстепенные члены – 0.01 < Di < 0.1.Отнесение входящих в состав биоценоза видов к доминантным группам только по их числовому обилию, как отмечал В.Ф. Ласточкин [1930, цит. по: Методика изучения.., 1975], было бы неправильным, так как более мелкие формы, как правило, наиболее многочисленны и всегда будут оказываться руководящими. Ряд исследователей [Руководство по гидробиологическому.., 1992] предлагают пользоваться для этого биомассой, т.е. весом на единицу площади дна, характеризующим массу живого органического вещества, которую образует вид, подставляя ее значения
Bi и Bs в формулу (4.4) того же индекса доминирования по В.Ф. Палию и А. Ковнацкому:Di
= 100 × pi × Bi / Bs.Но нетрудно увидеть, что вариационный размах натуральных значений биомассы (колеблющейся, в зависимости от размера животных, в сотни тысяч раз – от 1 мг до десятков и сотен грамм на 1 м
2) так же велик, как и для численности, в результате чего ранжирование видов по биомассе приводит к тому, что некоторые животные, например, моллюски, всегда будут иметь высокий приоритет, даже если на организменном уровне встречаются редко.Целенаправленные функциональные преобразования над значениями численности или биомассы в отдельности (например, логарифмирование или извлечение квадратного корня) могут несколько улучшить общую картину соотношения доминирования, поскольку модифицируют характер статистического распределения, приближая его к нормальному. Например, для рассматриваемой задачи Ф.Д. Мордухай-Болтовским [Методика изучения.., 1975] предложена более работоспособная форма индекса доминирования:
Di =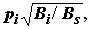
где
pi = mi / M – встречаемость вида i, mi – число проб, в которых был найден вид, M – общее число проб, Bi / Bs – удельная биомасса вида.Еще один интересный вариант видится нам в использовании различных функциональных комбинаций из трех основополагающих показателей (встречаемость, численность, биомасса), что дает возможность получить целый класс индексов, часть из которых в рамках поставленной
задачи могут оказаться состоятельными, несмещенными и эффективными оценками изучаемого явления. Примером такого обобщенного показателя является индекс плотности населения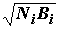 , введенный в гидробиологическую практику Л.А.Зенкевичем и В.А.Броцкой [1937] и широко использованный нами в математических расчетах части 3. Например, использование плотности населения в формуле (4.4)
, введенный в гидробиологическую практику Л.А.Зенкевичем и В.А.Броцкой [1937] и широко использованный нами в математических расчетах части 3. Например, использование плотности населения в формуле (4.4)
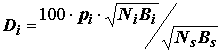
(4.5)
дает возможность выполнить ранжирование видов по доминантности с учетом обоих факторов – численности и биомассы.
Индексы доминантности, основанные на вероятности
pi , дают возможность ранжировать виды в пределах изучаемого объекта: сравнивать разные водоемы с использованием Di можно только в том достаточно редком случае, когда количество измерений М на каждом из них примерно одинаково [Тодераш, 1984].
Энтропийный подход к оценке биоразнообразия
Степень сложности биологических систем может рассматриваться также в аспекте эквитабельности компонентов видовой структуры. При этом под разнообразием (однородностью, выравненностью) обычно понимают то свойство [Левич, 1980], которое отличает, например, сообщество из 12 особей трех видов с распределением (4, 4, 4) от сообщества (10, 1, 1). Стремление представить характер такого распределения, заданного большим набором численностей, в компактном виде одним числом приводит к обобщенным индексам, форма выражения которых традиционно связывается с мерами количества информации.
Сделаем предварительно краткий экскурс в основные понятия теории информации.
Рассмотрим два независимых опыта a и b , таких что любые сведения об исходе одного опыта никак не меняют вероятностей исхода другого. Пусть опыт a
имеет r равновероятных исходов, а опыт b – l исходов. Рассмотрим сложный опыт a b , состоящий в одновременном выполнении обоих опытов. Степень неопределенности h(a b ) опыта a b равна сумме неопределенностей опытов a и b и, в соответствии с точкой зрения американского инженера-связиста Р. Хартли [Hartley, 1928, цит. по: А.М. Яглом, И.М. Яглом, 1973], может быть выражена следующим логарифмическим уравнением:h
(a b ) = h(a b ) + h(a b ) = log(r) + log(l).К. Шеннон [Shannon, 1947, рус. пер. 1963], развивая идеи Хартли, определил энтропию
опыта Н, как среднее значение неопределенности отдельных исходов:H
(a b ) = - (1/r) log(1/r) – (1/l) log(1/l)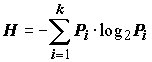 (4.6)
(4.6)
Энтропия (или неопределенность исхода) равна нулю, если вероятность одного из событий равна 1, и принимает максимальное значение в случае равновероятных исходов. Действительно, если известно, что в водоеме присутствует только один вид гидробионтов, то какая-либо неопределенность по его извлечению отсутствует (т.е. Н = 0). Неопределенность в предсказании результата отлова резко возрастает, если мы имеем в водоеме
k видов с одинаковой численностью. Важным для биологии свойством энтропии является то, что значительным числом исходов, суммарная вероятность которых мала, при подсчете энтропии можно пренебречь.Энтропию
H, как меру неопределенности, нельзя отождествлять с информацией I (как, например, напряженность электрического поля нельзя отождествлять с разностью потенциалов). Но количество информации об опыте b , содержащейся в опыте a , равноI
(a,b) = H(b) - Ha(b) ,(4.7)
где
Ha (b ) – условная энтропия опыта b после выполнения опыта a (т.е. снижение неопределенности b в результате выполнения a ). Для дальнейшего изложения приведем также известную из школьной комбинаторики формулу для числа перестановок из предметов k различных типов, общее число которых N = n1+n2+…+nk (авторство формулы маячит в глубине XVII в. между Б. Паскалем и Я. Бернулли):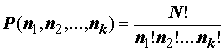 (4.8)
(4.8)
Опуская математические доказательства, приводимые А.М. Ягломом и И.М. Ягломом [1973], запишем равенство, в котором
H – энтропия: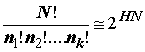
(4.9)
Трудно определить научный приоритет использования формулы (4.6) применительно к экосистемам. По сведениям И.И. Дедю [1990], Р. Мак-Артур уже в 1955 г. для оценки степени структурированности биоценозов использовал общее уравнение энтропии произвольной системы, опубликованное К. Шенноном в 1947 г. В 1957 г. Р. Маргалеф постулировал теоретическую концепцию, согласно которой разнообразие соответствует неопределенности (т.е. энтропии) при случайном выборе видов из сообщества [Макрушин, 1974а]. В результате этих работ большое распространение и повсеместное признание получил индекс Шеннона
Н, иногда называемый информационным индексом разнообразия К.Шеннона – У. Уивера.При расчете энтропии Н по Шеннону считается, что каждая проба – случайная выборка из сообщества, а соотношение видов в пробе отражает их реальное соотношение в природе. В качестве оценок вероятностей независимых событий р
i для формулы (4.6) могут быть использованы следующие апостериорные отношения:Чуть позже Р. Маргалеф [
Margalef, 1958], ссылаясь на формулу Л. Больцмана для энтропии изолированных термодинамических систем, предложил другое выражение для индекса разнообразия: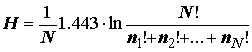 , (4.10)
, (4.10)
где
N! – факториальная величина всех исследуемых видов, ni! – факториал от числа особей каждого вида. Сопоставляя формулы (4.6) и (4.9), нетрудно увидеть, что (4.10) – просто иная форма расчета энтропии по Шеннону, не нашедшая, впрочем, широкого применения, поскольку, например, факториал от 34 составляет 3´ 1038 и является последним факториалом, вычисляемым на современном компьютере без использования специальных приемов.Использование индекса Шеннона Н в экологии связано с рядом его действительных и мнимых свойств, которые нам представляется интересным обсудить:
Для элиминации влияния числа видов
S Э. Пиелу [Pielou, 1966М, 1975] предложила индекс выравненности экологических сообществ Пиелу:H’
= H / Hmax = H/ log2 (1/S),который зависит только от равномерности распределения обилия по таксонам, поскольку представляет собой степень уклонения энтропии Н от ее максимального значения
Hmax = log2 (1/S) – см. кривую 1 на рис. 4.3.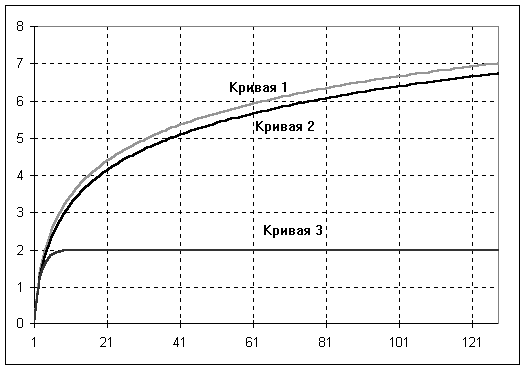
Рис. 4.3. Зависимость индекса Шеннона Н от числа видов
S
Очень простой метод оценки разнообразия предложен Р. Макинтошем [McIntoch, 1973; цит. по: Миркин и др., 1989] (индекс разнообразия Макинтоша
):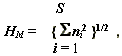
где
ni – оценка значимости каждого вида i (численность или биомасса), S – общее число видов. Возведение в квадрат значимости вида, естественно, "дает преимущество" в оценке разнообразия доминирующим видам. Еще раз подчеркнем, что этот индекс является прекрасной иллюстрацией “индексологии” – почему не в кубе? не в пятой степени? и вообще, какое отношение имеет представленная формулой Макинтоша среднеквадратическая численность к оценке биоразнообразия?...Другой подход к оценке разнообразия основан на подсчете количества возможных связей между внутренними элементами экосистемы, обеспечивающих ее единство и функционирование. Эти связи имеют смысл числа степеней свободы внутривидовых и межвидовых взаимодействий. В качестве меры связности может быть использован индекс разнообразия К. Гайни - Е. Симпсона [С.Gini, 1912М; Simpson, 1949; Миркин и др., 1989], больше известный как индекс Симпсона, который имеет вид:
C = S
(ni / N)2 или 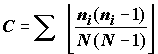 ,(4.11)
,(4.11)
где
ni – оценка значимости каждого вида (численность или биомасса), N – сумма оценок значимостей. Поскольку при возведении в квадрат малых отношений ni / N получаются очень малые величины, индекс Симпсона тем больше, чем сильнее доминирование одного или нескольких видов. Глубинный смысл индекса Симпсона выясняется, если принять во внимание, что знаменатель N (N-1)/2 означает число возможных пар особей независимо от их принадлежности к определенному таксону, а числитель n(n-1)/2 – число возможных пар особей одного таксона [Алимов, 2000].Р. Фишер с соавторами [Fischer et al., 1943М] нашел, что логарифмический ряд хорошо передает распределение видов по их численности, и предложил функцию a
как меру разнообразия:a m = (ln Nm - ln N1) / m
где
Nm – численность вида m в ряду видов, ранжированных по численности, N1 – численность первого вида с наивысшей численностью, m – порядковый номер вида в ряду 1, 2, 3, ..., m.Другим выражением для значимости вида
nr в ряду от наименее значимого (i = 1) к наиболее значимому (i = S) является формула Р. МакАртура [MacArthur, 1957]: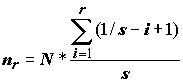 ,
,
где
N – сумма некоторых показателей обилия, по которым сравниваются виды, S – число видов.А. Корбет и К. Вильямс [Corbet, Williams, 1973; цит. по: Василевич, 1972] предложили уравнение, связывающее число видов
S с числом видов в биоценозе:S
= a * log(1 + N / a ) ,где log – Неперов логарифм, а параметр a предлагается как показатель разнообразия. Значение a возрастает по мере увеличения доли редких видов (с небольшой плотностью) в общем видовом пространстве биоценоза.
М. Хилл с соавт. [Hill et al., 1975] для оценки биоразнообразия предложил семейство средних степенных кривых, где
S – число видов, pi – оценки вероятности: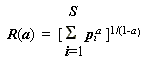 .(4.12)
.(4.12)
Легко показать [Розенберг c cоавт., 1999], что при разных значениях параметра а формулы (4.12) можно получить целый спектр индексов разнообразия, что свидетельствует об их "генетическом" родстве. Так,
R(0) = S, R(1) = f(H) – экспоненциальная версия индекса Шеннона, R(2) = 1 / C, где C – индекс Симпсона и т.д.| Дальше
|
Назад |
Начало |
Конец |
Список
|