| Выпуск 2
|
Конец |
Список
|
ПРАКТИКУМ ПО МАТЕМАТИЧЕСКИМ МЕТОДАМ В ЭКОЛОГИИ:
Выпуск 1
С.С. Крамаренко
МЕТОДЫ ОЦЕНКИ АБСОЛЮТНОЙ ПЛОТНОСТИ И ПРОСТРАНСТВЕННОЙ СТРУКТУРЫ ПОПУЛЯЦИИ
Введение
1. Методы оценки абсолютной плотности
2. Методы анализа пространственной структуры
2.1.2. Индекс дисперсии Соутвуда
2.1.3. Индекс Морисита
2.2.2. Метод Ивао
Введение
Определение численности объекта исследования оказывается первейшей задачей любого демэкологического и синэкологического исследований, поскольку она (верней, характер ее флуктуаций) теснейшим образом связана с различными аспектами жизнедеятельности организма. Численность популяции, по сути, является главной результирующей функцией, обусловленной размножением, смертностью, пространственным распределением, миграциями, социальными отношениями и т.д. организмов на фоне проявления различных экологических факторов.
Без данных о численности видов, которые входят в состав биоценоза, нельзя судить об их относительной роли в формировании стабильности данного сообщества, невозможно установить особенности развития биоценоза в пространстве и времени, выяснить характер динамики популяций отдельных видов.
Для решения перечисленных выше, а, кроме того, и многих других вопросов, нельзя ограничиваться только субъективной, например, глазомерной оценкой численности. Основная задача количественного учета организмов при проведении экологических исследований состоит в получении объективных и сопоставимых с результатами других исследований данных об обилии организмов данного вида в пределах определенной территории.
Наиболее сложной проблемой при определении показателей численности вида является определение точных границ популяции. Часто при проведении экологических исследований за границы популяций принимают резкие географические барьеры, непреодолимые для организмов, или административные границы. Последний подход менее приемлемый, но, тем не менее, широко распространен.
Одним из показателей обилия организмов является абсолютная плотность: наиболее вероятное число особей данного вида, которое приходится на единицу площади.
Наиболее распространенным и приемлемым методом определения абсолютной плотности для многих мелких и средних размеров седентарных животных и растений есть метод пробных площадок. Процедура определения искомого показателя состоит в следующем. Деревянную или металлическую квадратную рамку (лучше всего легкую и разборную) известной площади бросают через плечо, не глядя, и там где она упадет, отмечают всех особей данного вида (или нескольких видов одновременно), которые оказалось внутри рамки. Методически заранее решают, как быть с организмами, которые попали на границу рамки — включать их в анализ или не включать. Набор данных о численности особей в пределах всех пробных площадок, проанализированных в данный момент времени в данном месте их обитания, называется выборкой.
Площадь рамки зависит от величины объекта исследования и от его численности. Наиболее часто для почвенных беспозвоночных, бентосных организмов и растений используют квадратные рамки площадью 0,25 м2 или 1 м2. В некоторых случаях площадь рамки может быть 4 м2 или даже более.
Важнейшим методическим требованием при использовании данного метода является требование о случайном расположении рамок на территории исследуемого участка. Иногда предварительно строят карту исследуемого участка, разбивают его сеткой на квадраты, а потом с помощью генератора случайных чисел (данная функция имеется во многих микрокалькуляторах, а таблицы случайных чисел представлены в большинстве учебников по биометрии или математической статистике) определяют номера квадратов, в которых будут заложены пробные площадки. Приемлемо и расположение пробных площадок внутри однородного биотопа в шахматном порядке.
Приведем несколько основных условий, которые могут повлиять на результаты количественного учета численности организмов при использовании метода пробных площадок:
1. Чем большее будет заложено пробных площадок, тем надежнее (достовернее) будет полученный результат.
2. Чем большее будет площадь пробной площадки, тем более точны полученные данные.
3. Чем изменчивее явление (т.е. плотность организмов) и, соответственно, полученные показатели, тем большим должно быть количество наблюдений.
4. Массовые явления и грубые зависимости улавливаются уже при небольшом количестве выборок, и наоборот.
5. Конечная точность статистического результата в большей степени зависит от числа повторений, чем от педантичности проведения отдельного наблюдения. (Разумеется, необходимо строго придерживаться стандартности методики.)
6. Надежность экстраполяции (перенесение итогов выборочных наблюдений на всю территорию) тем выше, чем большая площадь была охвачена выборками, и чем большим было количество пробных площадок.
1. Методы оценки абсолютной плотности
Абсолютная плотность
(D) организмов на исследуемом однородном участке определяется как среднее арифметическое количества зарегистрированных в границах каждой пробной площадки (xi) организмов (данного вида):|
|
(1.1) |
где n — количество исследованных пробных площадок (i Î [1, n]).
Показателем, который характеризует точность получаемых результатов, является ошибка оценки плотности (SED), которая определяется через понятие ее вариансы (S2):
|
|
(1.2) |
|
|
(1.3) |
где
∑xi2 — сумма квадратов количеств организмов в любом из n пробных площадок, а (∑xі)2 — квадрат суммы этих величин. Достоверными считаются результаты, для которых отношение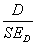 оказывается более или равно пяти, то есть величина ошибки должна составлять не более 20 % величины самого показателя.
оказывается более или равно пяти, то есть величина ошибки должна составлять не более 20 % величины самого показателя.
На основании этого можно определить необходимое (минимальное) количество пробных площадок так, чтобы полученные результаты были достоверными. Эта величина (nmin) определяется по формуле Эллиота (Elliot, 1971):
|
|
(1.4) |
Если численность организмов невелика в границах исследуемой территории, а самая территория имеет довольно большую площадь, тогда вместо площадок квадратной формы используются ленточные трансекты. Ленточная трансекта, по сути, та же пробная площадка (но большей площади), одна сторона которой намного больше другой (она представляет собой прямоугольник шириной от 0,5 до нескольких метров и длиной до нескольких сотен метров, в зависимости от размеров исследуемой территории). Методика определения количества особей, которые оказываются в пределах трансект и методы расчета оценки абсолютной плотности те же, что и для пробных площадок небольших размеров квадратной формы.
* * * * *
Для очень многих организмов тип распределения особей (или нор, гнезд и т.п.) в пространстве носит случайный характер (раздел 2). Математически это выражается в том, что число пробных площадок с 0, 1, 2, 3, ..., i и т.д. объектами задается рядом Пуассона:
|
|
(1.5) |
где n — общее количество исследованных пробных площадок; D — средняя плотность объектов на единицу территории.
Характерной особенностью распределения Пуассона есть то, что оно задается всего одним показателем. В нашем случае, это – оценка плотности популяции (или численности, которую легко пересчитать, зная плотность и число исследованных пробных площадок: N = D·n).
Если использовать прием оценки плотности, описанный выше, то пришлось бы подсчитывать количество организмов на каждой пробной площадке, а потом рассчитывать их среднее арифметическое и вариансу (для определения уровня точности оценки плотности популяции). При довольно высокой разреженности популяции (когда в пределах пробной площадки встречаются считанные объекты) для повышения уровня достоверности получаемых оценок плотности необходимо значительно увеличивать количество исследованных пробных площадок. С другой стороны, если объекты многочисленны (но обязательно распределены случайно), то приходится подолгу пересчитывать их в каждом квадрате.
Описанный ниже подход в значительной мере облегчает оценку плотности (и численности) особей (или других объектов), поскольку базируется не на непосредственном подсчете числа объектов, а на подсчете числа пробных площадок, в пределах которых был встречен хотя бы один организм (если плотность низка), или, наоборот, в пределах которых не встретилось ни одного организма (если плотность высока).
При случайном (пуассоновском) распределении объектов в границах исследованной территории, количество пробных площадок, в которых не будет найдено ни одного объекта, можно рассчитать по формуле:
|
|
(1.6) |
найден один объект:
|
|
(1.7) |
найдено два объекта:
|
|
(1.8) |
найдено три объекта:
|
|
(1.9) |
и т.д.
Если большинство пробных площадок содержат 1, 2, 3 и т.п. объектов, то нет необходимости подсчитывать их количество, достаточно подсчитать количество пробных площадок, в которых не было зарегистрировано ни одного объекта. Тогда количество площадок, которые содержат хотя бы один объект можно рассчитать по формуле:
|
|
(1.10) |
где n* — число площадок, которые содержат хотя бы один объект.
Отсюда оценка плотности популяции может быть рассчитана, как:
|
|
(1.11) |
(если подсчитывалось число пробных площадок, на которых не встречено ни одного объекта), или:
|
|
(1.12) |
(если подсчитывалось число пробных площадок, на которых был встречен хотя бы один объект).
Основным требованием при использовании данного метода является необходимость соответствия пространственного распределения организмов в анализируемой популяции закону Пуассона. Точность данного метода, естественно, повышается с увеличением количества пробных площадок.
Поскольку для распределения Пуассона среднее арифметическое равно его вариансе (то есть, D = S2), то ошибку оценки средней плотности можно найти по формуле:
|
|
(1.13) |
Однако в некоторых случаях вместо n пробных площадок площадью S можно проанализировать наличие объектов (животных, гнезд, нор и т.п.) на одной пробной площадке общей площадью S1 = n·S, то есть, вместо большого числа небольших пробных площадок использовать одну ленточную трансекту (см. выше).
Количество объектов, зарегистрированных в пределах трансекты (N), отнесенное к ее площади (S1), даст оценку абсолютной плотности (D):
|
|
(1.14) |
Если характер распределения объектов на исследованной территории случайный, то оценку ошибки показателя плотности популяции можно получить следующим образом (на основании особенностей распределения Пуассона):
|
|
(1.15) |
При большой численности объектов, зарегистрированных на территории одной трансекты (или на нескольких десятках пробных площадок), доверительные интервалы оценки плотности рассчитываются по формуле: D + 1,96× SED.
Но поскольку при низких значениях среднего арифметического (то есть, D, а соответственно и N) распределение Пуассона становится резко асимметричным, то при небольших значениях плотности, полученных на основании приведенной выше методики, доверительный интервал строится иначе.
В этом случае доверительный интервал для общего числа объектов (особей, нор, гнезд и т.п.), зарегистрированных на исследуемой территории (то есть N), рассчитывается по формулам:
|
N н = 0,5× c 21 (df1 = 2× N; p = a/2), |
(1.16) |
|
N в = 0,5× c 22 (df2= 2× N + 2; p = 1 - a/2), |
(1.17) |
где Nн и Nв — нижняя и верхняя границы показателя; c 2 — табличное значение критерия Хи-квадрат Пирсона с заданным числом степеней свободы (df),
a - уровень значимости.При необходимости, для оценки уровня достоверности различий между двумя показателями плотности (D1 и D2), полученными с использованием данной методики, например для двух разных участков, можно использовать u-критерий:
|
|
(1.18) |
а для проверки достоверности различий между двумя оценками численности (N1 и N2) объектов в пределах ленточной трансекты его следующую модификацию:
|
|
(1.19) |
где n1 и n2 ¾ количество пробных площадок, которые были использованы при анализе плотности на двух территориях, между которыми производится сравнение.
Рассчитанное значение u сравнивается с критическим значением (при уровне значимости
a = 0,05), которое составляет u = 1,65. Если u > ua = 0,05, то нуль-гипотеза отвергается, и наличие достоверных различий между двумя оценками считается доказанным.Но при низких значениях оценок, если N < 50, аппроксимация распределения Пуассона нормальным уже невозможна, и тогда необходимо использовать другой подход.
Он предусматривает использование или равного количества пробных площадок в обоих исследуемых местах обитания, или транскет равных размеров.
В этом случае, прежде всего, из двух оценок численности выбирается меньшая (N1). В таблице 1.1 против этого значения находится критическое значение другого (большего) значения (Nтабл). Тогда, если большее значение оценки численности (N2) не превышает критического, то наличие достоверных различий между оценками N1 и N2 признается не доказанным и, соответственно, N1 = N2. Если же N2 > Nтабл, то оценки численности, которые сравниваются различаются достоверно между собою на уровне значимости p < 0,05.
Таблица 1.1
Критерий различий двух оценок численности
|
N1 |
Nтабл |
N1 |
Nтабл |
N1 |
Nтабл |
N1 |
Nтабл |
N1 |
Nтабл |
|
1 |
6 |
11 |
21 |
21 |
34 |
31 |
46 |
41 |
58 |
|
2 |
8 |
12 |
22 |
22 |
35 |
32 |
47 |
42 |
59 |
|
3 |
9 |
12 |
23 |
23 |
36 |
33 |
48 |
43 |
60 |
|
4 |
11 |
14 |
25 |
24 |
37 |
34 |
49 |
44 |
61 |
|
5 |
12 |
15 |
26 |
25 |
39 |
35 |
51 |
45 |
63 |
|
6 |
14 |
16 |
27 |
26 |
40 |
36 |
52 |
46 |
64 |
|
7 |
15 |
17 |
28 |
27 |
41 |
37 |
53 |
47 |
65 |
|
8 |
17 |
18 |
30 |
28 |
42 |
38 |
54 |
48 |
66 |
|
9 |
18 |
19 |
31 |
29 |
43 |
39 |
55 |
49 |
67 |
|
10 |
19 |
20 |
32 |
30 |
45 |
40 |
57 |
50 |
68 |
2. Методы анализа пространственной структуры
Не менее важной экологической характеристикой исследуемой популяции является характер пространственного распределения особей.
Если организмы не склонны к проявлению каких-либо устойчивых (отрицательных или положительных) взаимодействий, они будут распределены в пространстве случайным образом. Если же отношения между особями в своей основе антагонистические (отрицательные), между ними действуют силы отталкивания, поэтому, в относительно однородной среде обитания их пространственное размещение окажется близким к равномерному. И, наконец, если у организмов имеет место стремление к положительным взаимодействиям, их размещение в однородной среде может быть групповым. Кроме того, групповой тип распределения организмов в пространстве может быть связан с неравномерностью (гетерогенностью) самих абиотических или биотических факторов внешней среды.
Пространственная структура популяции может претерпевать изменения при разных уровнях численности. Например, при повышении численности свыше какого-либо определенного уровня равномерное или групповое распределение может сменяться случайным. Таким образом, в общем случае перенаселение ведет к уменьшению уровня организации.
2.1. Методы оценки характера пространственного распределения, основанные на однократном учете
Все методы определения характера пространственного распределения особей основаны на одном важном свойстве случайного распределения - для этого типа распределения среднее арифметическое равняется его вариансе. Таким образом, если мы определим среднюю плотность организмов в исследуемой популяции методом пробных площадок или трансект (см. раздел 1) и рассчитаем вариансу этого показателя, то при D = S2 распределение особей окажется случайным, при D > S2 — равномерным, а при D < S2 — групповым.
Необходимо указать, что при изменении масштаба пробной площадки, тип распределения особей в пространстве может изменяться. Поэтому обязательно необходимо придерживаться однотипности приемов анализа популяции (или популяций) как в пространстве, так и во времени для получения сравниваемых показателей.
2.1.1. Индекс Одума
Наиболее простым методом определения характера пространственного распределения объектов является использование индекса Одума (IO) (Одум, 1986):
|
|
(2.1) |
Если статистически доказано, что IO < 1, то особи распределены в популяции равномерно, если IO > 1 — группами и, наконец, если IO = 1, то особи распределенные случайным образом.
Индекс Одума характеризует тип пространственного распределения в каждый момент сбора и, следовательно, может быть использован для анализа изменения пространственной структуры популяции во времени.
Уровень значимости индекса Одума можно оценить, сравнивая рассчитанное значение IO с табличным значением F-критерия Фишера-Снедекора с числом степеней свободы df1 = df2 = n – 1 (Таблица 2.1).
Таблица 2.1
Табличные значения F-критерия Фишера-Снедекора для df1 = df2 и
a = 0,05|
df |
F |
df |
F |
df |
F |
df |
F |
df |
F |
|
1 |
161,4 |
11 |
2,82 |
21 |
2,08 |
31 |
1,82 |
45 |
1,64 |
|
2 |
19,0 |
12 |
2,69 |
22 |
2,05 |
32 |
1,80 |
50 |
1,60 |
|
3 |
9,28 |
13 |
2,58 |
23 |
2,01 |
33 |
1,79 |
55 |
1,56 |
|
4 |
6,39 |
14 |
2,48 |
24 |
1,98 |
34 |
1,77 |
60 |
1,53 |
|
5 |
5,05 |
15 |
2,40 |
25 |
1,96 |
35 |
1,76 |
65 |
1,51 |
|
6 |
4,28 |
16 |
2,33 |
26 |
1,93 |
36 |
1,74 |
70 |
1,49 |
|
7 |
3,79 |
17 |
2,27 |
27 |
1,90 |
37 |
1,73 |
75 |
1,47 |
|
8 |
3,44 |
18 |
2,22 |
28 |
1,88 |
38 |
1,72 |
80 |
1,45 |
|
9 |
3,18 |
19 |
2,17 |
29 |
1,86 |
39 |
1,70 |
90 |
1,42 |
|
10 |
2,97 |
20 |
2,12 |
30 |
1,84 |
40 |
1,69 |
100 |
1,39 |
В тех случаях, если D > S2 , рассчитывается обратная величина — D/ S2, которая затем сравнивается с табличным значением F-критерия Фишера-Снедекора с числом степеней свободы df1 = df2 = n - 1. Если рассчитанное значение превышает табличное, то распределение носит равномерный характер.
При большом количестве числа степеней свободы уровень значимости F-критерия Фишера-Снедекора может быть также оценен с помощью u-преобразования:
|
|
(2.2) |
где k = 2 : (9× df).
Если рассчитанное значение u превышает критическое u
a=0,05 = 1,65, делается вывод, что рассчитанное значение F-критерия Фишера-Снедекора достоверно превышает соответствующее табличное для соответствующего уровня значимости.2.1.2. Индекс дисперсии Соутвуда
Некоторой модификацией индекса, предложенного Одумом, есть индекс дисперсии Соутвуда (Id) (Southwood, 1978). С его помощью можно оценить достоверность отличия анализируемого распределения от равномерного:
|
|
(2.3) |
Для оценки уровня значимости, рассчитанное значение данного показателя сравнивают с табличными значениями критерия Хи-квадрат Пирсона для числа степеней свободы df = n - 1 (то есть число пробных площадок минус единица).
При использовании индекса Соутвуда может встретиться ситуация, когда значение числа степеней свободы будет превышать табличные значения критерия Хи-квадрат Пирсона. В этом случае, достоверность отличия полученного значения индекса Соутвуда от табличного определяется с помощью u-преобразования:
|
|
(2.4) |
В том случае, если полученное значение u превышает критическое u
a=0,05 = 1,65, то считается доказанным, что рассчитанное значение индекса Соутвуда превышает табличное значение критерия Хи-квадрат Пирсона и распределение объектов достоверно отклоняется от равномерного.Удобство u-преобразования состоит в том, что оно может быть использовано при любом числе степеней свободы.
2.1.3. Индекс Морисита
Один из недостатков описанных выше способов оценки характера пространственного распределения организмов в популяции состоит в допущении того, что выборка намного меньше, чем образованные особями скопления. Очевидный выход, который разрешает нивелировать такое допущение, заключается в использовании большого числа небольших выборок (пробных площадок).
Методом, который лишен этого недостатка, является оценка индекса Морисита (Morisita, 1959). На результаты использования данного метода не оказывают влияния ни размеры пробной площадки, ни размеры выборки (при условии, что она не слишком большая).
Формула для оценки индекса Морисита следующая:
|
|
(2.5) |
где n — общее количество использованных пробных площадок; f — количество площадок, которые содержат Ni особей; N — общее количество особей на всех площадках (то есть, N =
S Ni).Индекс Морисита равняется Id = 1 при случайном типе распределения особей (объектов) в пространстве, Id < 1 - при равномерном, и Id > 1 - при групповом.
2.2. Методы оценки характера пространственного распределения, основанные на многократных учетах
Все предложенные выше методы характеристики пространственного распределения особей (объектов) в популяции используются при одномоментном анализе, то есть дают характеристику типа распределения при одноразовом сборе материала. Однако, как мы уже указывали выше, характер пространственного распределения особей в популяции может существенным образом изменяться во времени. Два предложенных ниже метода характеризуют типичный для данной популяции характер пространственного распределения в течение продолжительного периода времени, а их расчет базируется на наборе пар значений средней плотности и их варианс, полученных для отдельных серий одномоментных сборов объектов.
2.2.1. Уравнение Тейлора
Средняя плотность особей в популяции чаще всего испытывает значительные временные колебания. Поскольку при увеличении средней плотности возрастает также оценка вариансы плотности, то в таких случаях зависимость между данными оценками может быть описана выражением:
|
|
(2.6) |
где k — показатель, который характеризует изменчивость оценок средней плотности в разные моменты времени.
В этом случае распределение особей в пространстве хорошо аппроксимируется отрицательным биномиальным распределением. Данное распределение выводится из распределения Пуассона, но базируется на предположении, что среднее значение не фиксировано, а изменяется случайно. Такое распределение может быть применимо к популяциям, в которых организмы формируют скопления, причем последние распределены случайным образом.
Если показатель k® +¥ , то отрицательное биномиальное распределение приближается к распределению Пуассона, то есть распределение особей в популяции (или их скоплений) имеет случайный характер, если же k<<+¥ , то особи в популяции собраны в более или менее многочисленные скопления (Макфедьен, 1965).
В таких случаях характер пространственного распределения особей в популяции может быть оценен, используя параметры уравнения Тейлора (Taylor, 1961), которое описывает зависимость между показателями варианс и средней плотности, оцененных для исследуемой популяции во время нескольких последовательных посещений (период исследования, как правило, несколько лет, а общее количество посещений за это время — 20-30):
|
|
(2.7) |
Если значение коэффициента b уравнения Тейлора близко к 1, то мы имеем дело со случайным распределением, если достоверно превышает 1, то распределение особей имеет групповой характер.
Использование уравнения Тейлора основано на той особенности отрицательного биномиального распределения, что оно с высокой степенью точности может быть аппроксимировано показательной зависимостью, то есть:
|
|
(2.8) |
При этом если показатель отрицательного биномиального распределения k® +¥ (то есть, распределение особей носит случайный характер), показатель степени уравнения Тейлора b ® 1.
Важным достоинством данного метода является то, что величина параметра b совершенно не зависит ни от числа пробных площадок, ни от общего числа организмов в них. Этот параметр является важной характеристикой разных видов (или популяций), поскольку его значение остается относительно постоянным во времени.
Проверка статистического вывода о равенстве полученного значения параметра b единице (или достоверного превышения ее) производится, используя t-критерий Стьюдента:
|
|
(2.9) |
где SEb — ошибка параметра b. Полученное значение t-критерия Стьюдента сравнивают с табличным значением для df = n – 2.
2.2.2. Метод Ивао
С другой стороны, необходимо также учитывать тот факт, что структура популяции может быть очень сложной, например, особи могут образовывать скопления (разной величины), которые в свою очередь также могут быть распределены случайно, равномерно или группами второго порядка. Метод, который позволяет учитывать такой характер пространственной структуры популяции, это – метод Ивао (Iwao, 1968).
Он также основан на отношении вариансы к среднему значению плотности, а характер пространственного распределения особей в популяции характеризуется параметрами
α и β линейной зависимости:|
|
(2.10) |
где
|
|
(2.11) |
Для распределения одиночных особей значение коэффициента
α ≤ 0, а для группового распределения – α > 0; если же особи или колонии распределены случайным образом, то значение коэффициента β ≤ 1, а при групповом распределении – β > 1. Уровень значимости отличия рассчитанных оценок параметров α и β от граничных определяется с помощью t-критерия Стьюдента с учетом соответствующих величин ошибок (см. выше).
ЛИТЕРАТУРА
Макфедьен Э
. Экология животных: Цели и методы. Пер. с англ. — М.: Мир, 1965. — 376 с.Одум Ю. Экология: В 2-х т. Пер. с англ. — М.: Мир, 1986.
Elliot J.M. Some methods for the statistical analysis of samples of benthic invertebrates // Freshwater biol.Assoc. — 1971. — 25. — P.1-144.
Iwao S. A new regression method for analyzing the aggregation pattern of animal populations // Res.Populat.Ecol. — 1968. — 10, 1. — P.1-20.
Morisita M. Measuring the dispersion of individuals and analysis of the distributional patterns // Mem.Fac.Sci.Kyushu Univ. — 1959. — Ser. E 2. — P.215-235.
Soutwood T.R.E. Ecological methods. — London: Chapman & Hall, 1978. — 524 p.
Taylor L.R. Aggregation, variance and the mean // Nature (London). — 1961. — 189. — P.732-735.
| Выпуск 2
|
Начало |
Список
|
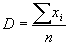
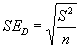
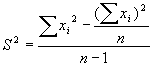
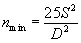
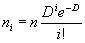
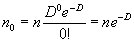
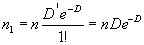
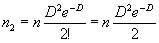
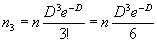
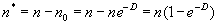
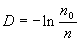
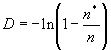
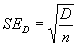
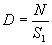
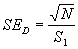
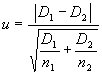
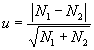
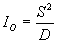
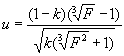 ,
,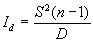
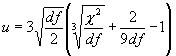 .
.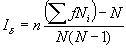
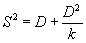
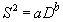
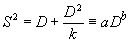 .
.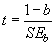
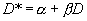 ,
,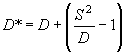 .
.