| Дальше
|
Назад |
Начало |
Конец |
Список
|
4.8. Элементы продукционной гидробиологии
Мем № 25
: “Биологическая структура является открытой нелинейной системой, которая препятствует своему разрушению за счет способности к самоорганизации. Фактически, жизнь есть не что иное, как система по понижению собственной энтропии за счет повышения энтропии окружающей среды. Но расплатой за устойчивость живой материи является ее зависимость от поступления энергии извне, как необходимого условия существования неравновесной биосистемы. Если способность системы к самоподдержанию своей структуры ослабевает, в том числе из-за неадекватного энергетического восполнения, то ее элементы становятся менее организованными и различия между ними постепенно нивелируются” С.Б. Пашутин [URL].Научное формирование развернутых представлений о механизмах трофических взаимосвязей и динамике биотического круговорота в водных экосистемах связано с развитием представлений о балансе вещества и энергии в водоеме, основные методологические концепции которых разработаны П. Бойзен-Иенсеном [Boysen-Jensen,1919], Л.А. Зенкевичем [1934], Г.Г. Винбергом [1936], В.С. Ивлевым [1938], Р. Линдеманном [Lindeman, 1942], А.Ф. Алимовым [1982, 1987, 1988] и др. Существуют многочисленные литературные источники [Шмальгаузен, 1935; Зенкевич, 19
51; Ивлев, 1955; Винберг, 1960, 1979а,б; Россолимо, 1964; Хатчинсон, 1969; Бульон, 1983, 1994; Заика, 1983; Гутельмахер, 1986; Алимов, 1989; Голубков, 2000], посвященные теоретическому обоснованию и принципам расчета функционально-энергетических характеристик различных групп водных организмов. В результате структурного анализа и математического моделирования биотических балансов к настоящему времени установлена и количественно выражена генеральная схема материально-энергетических потоков в разных по типу и характеру водных биоценозов. Отсылая заинтересованного читателя к фундаментальным трудам по этой тематике, в настоящем разделе мы ограничимся общей схемой расчета продукционно-деструкционных показателей на примере зообентоса. Крупные ученые - основоположники продукционной биологии

Процессы конструктивного и энергетического обмена в водной экосистеме основаны на тесно и закономерно взаимосвязанных механизмах биогеохимического круговорота, подчиняющегося общим законам сохранения материи Лавуазье–Ломоносова. Это дает возможность представить функционирование сообщества гидробионтов на любом организационном уровне (организм, популяция, биоценоз или водоем в целом) в виде балансовой модели: формальных структурных элементов-преобразователей, связанных материальными потоками и осуществляющих трансформацию вещества и энергии. Для каждого блока модели существует два вида потоков:
Для корректно сводимого материального баланса необходимы следующие условия:
Биотический баланс может быть выражен в абсолютных значениях (например, для всего водоема в целом), либо пересчитан на единицу наиболее важной ресурсной или итоговой составляющей: 1 джоуль ассимилированной солнечной энергии, 1 м
2 площади дна или 1 тонну образовавшейся биомассы. В отличие от материальных балансов искусственных технологических процессов, биотический баланс природной экосистемы в принципе не может быть стационарным, поэтому важнейшей его определяющей составляющей является период времени, за который он был составлен.Структура основных материальных потоков биотического баланса представлена на рис. 4.7, где объем поступления вещества с потребленной пищей (рацион) обозначен как С, количество не усвоенной пищи –
F, ассимиляция пищи гидробионтами – A:A = C - F
. (4.36)
(4.36)
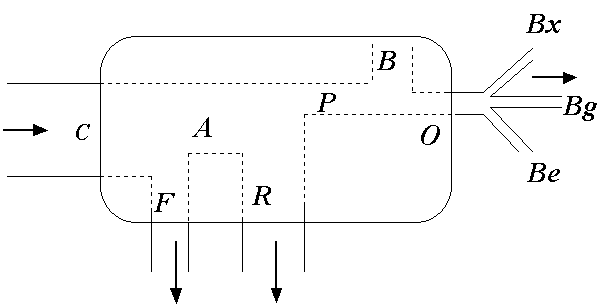
Рис. 4.7. Структура основных потоков обмена веществом и энергией с внешней средой организма или популяции (С – потребление или рацион, А – ассимиляция,F – не усвоенная пища, R – траты на обмен, О – отторгаемая продукция, Bg – половые продукты, Bx – экскреты, Be – экзувии, B – прирост биомассы)
Обмен веществ, связанный с жизнедеятельностью, сопровождается выполнением внешней и внутренней работы, сжиганием части ассимилированной пищи и, в конечном итоге, преобразованием энергии сгорания в тепловую. Интенсивность этого процесса трат на обмен обозначена как
R. Разность между потоками энергии, поступающей в организм с ассимилированной пищей, и потоком рассеиваемой тепловой энергией в результате трат на обмен представляет собой продукционный поток энергии P :P = A - R
.(4.37)
В терминах энергии процесс продуцирования можно определить как суммарный процесс накопления биомассы
B организмами и их популяциями в виде энергосодержащей соматической ткани, а также в виде отторгаемой продукции O, включающей генеративные (половые) продукты Bg, экскреты Bx и другие отчуждаемые продукты роста Be (экзувии, слизь,метаболиты).Уровень особи
Рассмотрим вкратце систему понятий, предельно четко сформулированных В.В. Меншуткиным [1971] и используемых для построения биотических балансов популяций и сообществ водных животных. В качестве первичного (неделимого) элемента модели принимается особь, существование которой в каждый момент времени определяется следующими основными компонентами: t – возраст особи, т.е. время, прошедшее с момента начала самостоятельного развития яйца или икринки; w – масса тела особи; k – калорийность тела особи (положительная переменная, имеющая размерность энергии, отнесенной к единице массы); с – реальный рацион, равный количеству пищи, потребляемой особью в единицу времени. Тогда рост особи – это изменение массы тела в единицу времени:
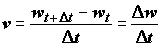 . (4.38)
. (4.38)
Энергия ассимилированной особью пищи в единицу времени (
aI) равна сумме трат на обмен r, изменения энергосодержания тела особи и энтальпии генеративных продуктов: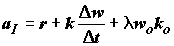 , (4.39)
, (4.39)
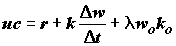 . (4.40)
. (4.40)
Важным показателем биоэнергетики особи является коэффициент использования ассимилированной энергии на рост (
k2), равный приросту массы тела на единицу массы усвоенной пищи с учетом их калорийности: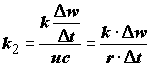 . (4.41)
. (4.41)
Коэффициент
k2, как показали работы В.С. Ивлева, закономерно снижается с увеличением возраста животного, в следствии того, что генеративный (экзувиальный) рост начинает превалировать над соматической составляющей.По литературным данным [Алимов, Финогенова, 1976] усвояемость пищи
u для всех нехищных животных, за исключением олигохет, может быть принята равной 0.6, для олигохет – 0.5, для хищных животных – 0.8. Средние значения калорийности k обычно считают равными 19.2 кДж/г сухого вещества для водорослей и макрофитов, а для водных животных – 23.01 кДж/г. Естественно, что калорийность организмов будет различаться в зависимости от того, рассчитывается ли она для сухой, сырой массы или органического вещества тела животных. Так энергетический эквивалент сырой массы донных организмов зообентоса варьируется в достаточно широких пределах от 4.18 кДж/г для Oligochaeta до 1.0 кДж/г для Dreissenidae (c раковиной).В практике биоэнергетических исследований последних десятилетий канонизировано применение степенных уравнений типа
Y = a Xk , (4.42)
которые, как считают многие авторы (см. [Винберг, 1976]), достаточно хорошо описывают параболические зависимости, постулируемые для расчета большинства компонентов материального баланса особи:
В частности, траты на обмен у донных животных при нормальных условиях обычно рассчитывают по уравнениям потребления кислорода животными в процессе дыхания в зависимости от массы тела, имеющим вид для:
ракообразных
олигохет
двустворчатых моллюсков ![]()
где
qo, мгО/ч – скорость обмена при t = 200C; w, г – масса тела особи. Чтобы перейти к тратам на обмен, выраженным в количестве диссипативной энергии ro, расходуемой особью на поддержание своей жизнедеятельности, необходимо количество потребленного кислорода qo умножить на оксикалорийный коэффициент 14.23 Дж/мгО. Ускоряющее влияние температуры среды на различные физиологические процессы, в частности, на интенсивность газообмена пойкилотермных животных, учитывается с использованием температурных поправок b t (t) в уравнении r = ro b t (t), которые могут быть взяты по “нормальной кривой” Крога [Винберг, 1956, 1983], либо с использованием коэффициентa Вант-Гоффа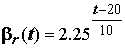 , коэффициента термолабильности Б.М.Медникова и проч.
, коэффициента термолабильности Б.М.Медникова и проч.
Наш комментарий:
Теоретически обосновывая канонизацию уравнения (4.42), Г.Г. Винберг [1956] и Н.П. Макарова [1975] связывают его с уравнением скорости анаболизма Л. фон Берталанфи [Bertalanffi, 1964]. Не отрицая известной адекватности параболической зависимости для описания широкого класса процессов биоэнергетики, мы считаем малопродуктивной догмой идею "универсализма" не совсем удобного в точных расчетах степенного уравнения Y = a Xk. Можно привести целый ряд иных более удобных математических формул, которые будут описывать те же экспериментальные данные не с меньшей степенью адекватности. Кроме этого, определенной методологической ошибкой статистической обработки [Умнов, 1976] является широко практикуемая линеаризация степенного уравнения путем логарифмирования в уравнение регрессии вида: lg Y = lg a + k lg x . Как будет показано ниже в главе 5, коэффициенты этих обоих уравнений, оптимальные относительно суммы квадратов невязок, далеко не идентичны между собой.Группа особей и популяция
Вторым уровнем системы модельных уравнений материально-энергетического баланса является группа (когорта) – множество особей, параметры состояния которых одинаковы или находятся в некоторых заданных пределах. Например, множество особей, находящихся на одинаковой стадии развития, образуют стадийные группы. По иному определяющему признаку могут быть скомплектованы весовые группы, возрастные группы и т.д. Состояние группы Ii Î
G определяется ее мгновенной численностью Nt и статистической функцией распределения всех регистрируемых показателей функционирования отдельных организмов. В большинстве балансов эти распределения достаточно описывать средними значениями: средний вес 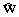 , средняя калорийность
, средняя калорийность 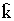 или средний возраст особи
или средний возраст особи 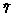 в группе.
в группе.
Суммарный вес особей данной группы составляет ее биомассу:
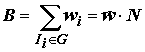 . (4.43)
. (4.43)
По аналогичным формулам могут быть рассчитаны: С – суммарный реальный рацион всех особей группы,
R – суммарные траты на обмен; U – суммарное количество несъеденной и непереваренной пищи; J – суммарная масса генеративных продуктов и метаболитов. Состав групп всегда нестационарен во времени и определяется динамикой смертности, а также процессами перехода особей из одной группы в другую. Например, изменение численности особей, входящих в стадийную группу Gi, описывается следующим соотношением: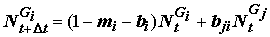 , (4.44)
, (4.44)
где
mi – коэффициент смертности особей, выловленных или съеденных хищниками за период D t, bi – коэффициент выбытия особей на следующую стадию, связанный с продолжительностью этапов развития; bji – коэффициенты поступления особей из других групп.Третий уровень иерархии баланса составляют популяции – множество особей или групп особей, обладающих свойством самовоспроизведения. Большинство характеристик и свойств группы особей можно перенести и на популяцию. Так появляется понятие о численности популяции
N, ее биомассе B и тратах на обмен R, как суммы показателей по всем возрастным группам, составляющим популяцию, а также среднем возрасте и среднем весе особи , которые рассчитываются как средневзвешенные значения по обычным статистическим формулам. Однако понятия, связанные с процессом перехода особей из одной группы в другую, не могут иметь смысла для популяции как для системы, замкнутой в отношении операций перехода и размножения. Кроме диссипативных потерь R и метаболитов, материально-энергетический выход из популяции P’ осуществляется только через биомассу погибших в единицу времени особей. Отношения размножения популяции заключаются в приросте численности особей в группах, имеющих соответственный стадийно-возрастной статус.
Специфической характеристикой популяции является понятие “продукции”. Под продукцией популяции понимается количество органического вещества, создаваемого всеми особями популяции в виде прироста массы их тела и выделения половых продуктов за единицу времени [Boysen-Jensen,1919; Винберг, 1936; Заика, 1983]. В энергетическом выражении продукция определяется как
 ,(4.45)
,(4.45)
где D
wi – прирост массы тела особей i-й группы популяции, независимо от того, остались ли они в живых, или погибли в течение временного шага системы D t; n – число групп в популяции. В тех же терминах энергетический выход популяции можно представить как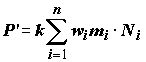 ,
,
где
mi – коэффициент смертности особей.Таким образом, популяцию можно определить, как материальный преобразователь, на вход которого подается вещество или энергия в виде пищи, а на выходе образуется органическое вещество иного уровня организации, которое является энергетической базой для популяций других трофических уровней. В стационарном случае, т.е. когда нет изменения биомассы популяции во времени (D
B = 0), продукция популяции равна ее энергетическому выходу и P = P’.В общем случае энергетический баланс популяции можно записать в виде
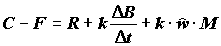 ;
; 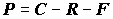 ;
; 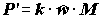 , (4.46)
, (4.46)
где
C – рацион; F – неусвоенная пища; M – количество особей, выбывших из популяции и использованных в пищу на других трофических уровнях.Компоненты биотического баланса популяции по сложившейся традиции используются для получения различных коэффициентов, которым зачастую придается особо важный, почти мистический смысл. Наиболее широко цитируемым является показатель интенсивности продукционного процесса (“удельная продукция” по В.Е. Заике [1983]) или отношение продукции популяции к ее средней биомассе, называемое также коэффициентом
P/B: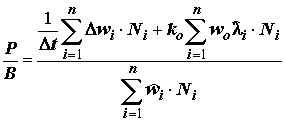 . (4.47)
. (4.47)
Численные значения величины продукции, а, следовательно, и коэффициента
P/B зависят от выбора единицы времени (суточные, недельные, месячные, годовые и прочие значения P/B). Для сравнения не абсолютной, а относительной интенсивности продукционного процесса иногда относят коэффициент P/B к средней продолжительности жизни особи как к естественному масштабу времени популяции. Однако до сих пор нет единого мнения (может – и не должно быть?) относительно того, какие величины P/B характерны для тех или иных объектов или условий их функционирования.Другой функциональной характеристикой сообществ организмов является соотношение Шредингера
P/R между продукцией и диссипативной энергией, рассеиваемой организмами в процессах обмена. А.Ф. Алимовым [1989] было показано, что для широкого класса популяций донных животных это соотношение за год или вегетативный сезон статистически постоянно и составляет 1.45 кДж/м2.Приведенный выше коэффициент
k2 (4.41) для особи сохранят свой смысл и для популяции: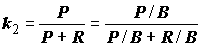 ,(4.48)
,(4.48)
однако его кумулятивные значения за достаточно большой промежуток времени не соответствуют законам онтогенетического развития отдельных особей, а зависят от динамики размерно-возрастного состава популяций, косвенно определяемой условиями внешней среды (трофические условия, температура и проч.) В любом случае, эффективность продукции, т.е.
k2 популяции, оказывается тем выше, чем больше в популяции доля особей младшего возраста, которым свойственны более высокие величины k2. Если принять во внимание постоянство отношения Шредингера (P/R = 1.45 кДж/м2 за сезон), то легко показать, что для большинства видов зообентоса среднесезонное значение k2 = 0.26. Это соотношение позволяет оценить приближенно продукцию популяции за вегетационный сезон по рассчитанным тратам на обмен (так называемый, "физиологический" метод).Балансы трофических уровней и биоценозов
Множество популяций, характерных сходными кормовыми связями, образует трофический уровень. Схематически трофо-динамическое направление в гидробиологии, основные положения которого разработаны Р. Линдеманом [Lindeman, 1942] и Дж. Хатчинсоном [1969] (см. раздел 2.2), сводится к следующему.
Первичное автохтонное органическое вещество образуется на первом трофическом уровне в ходе фотосинтеза зелеными растениями или хемосинтеза бактериями. "Пищей" в данном случае является аккумулированная солнечная энергия и аллохтонные органические вещества, поступающие в водоем извне с площади водосбора. В дальнейшем происходит поэтапная трансформация материи, синтезированной фитопланктонерами на первом уровне: на последующих трофических звеньях животные высшего уровня используют запас материи нижних уровней для построения своего тела. Каждый из выделенных уровней характеризуется своими компонентами биотического баланса: рационом С, ассимиляцией А, деструкцией или тратами на обмен R, продукцией P и массопереносом с уровня на уровень P’. Принято строго различать (по всей видимости, не из общетеоретических соображений, а из-за различий в технике расчетов) первичную продукцию, или продукцию автотрофов, и вторичную продукцию, или продукцию консументов-гетеротрофов.
Концепция трофических уровней позволила рассматривать процессы потребления, продуцирования, деструкции в сообществах во всей их взаимосвязи. Однако застывшая детерминированность трофических цепей не свойственна реальным экосистемам. Во многих случаях особи одного и того же вида на разных стадиях развития или при изменении внешних условий могут легко переходить с одной пищи на другую, а также менять тип и способ питания. Более строгое представление о трофических взаимодействиях дает односвязная трофическая сеть, т.е. более разветвленная по сравнению с трофическими уровнями система трофических связей, имеющая следующий формализм.
Пусть на множестве популяций (или групп особей) задан граф трофических связей Г(S), где два любых узла могут быть связаны ребром, направленным от хищника к жертве. Каждому ребру поставлена в соответствие трофическая ценность j-й жертвы по отношению к i-му хищнику как априорная вероятность ее поедания этим хищником. Тогда под биотическими сообществами можно понимать связные компоненты этого графа – фрагменты узлов сети, внутренне неразделимые, но изолированные друг от друга на некотором статистическом уровне значимости. Множество популяций биотического сообщества, составляющих изолированный фрагмент трофической сети, вместе с замкнутыми циклами биогенных элементов и компонентами неживой природы образуют экосистему или биогеоценоз. Из данного определения следует, что экосистема в отличие от сообщества обязательно включает в себя автотрофные элементы, т.к. в противном случае невозможно получение замкнутых биогеохимических циклов.
Определение продукции биоценозов – один из наиболее сложных и неоднозначно трактуемых вопросов продукционной гидробиологии и экологии в целом. С общетеоретических позиций продукция экосистемы (Ре) представляет собой разность между первичной продукцией экосистемы (Рр) и суммарными тратами на обменные процессы у всех гидробионтов системы (Re) [Алимов, 1987]: Ре = Рр—Re. В тех случаях, когда Re>Pp, существование биотической части экосистемы возможно только за счет поступления органических веществ извне.
Схема биотического баланса водных экосистем, впервые предложенная и составленная Г.Г. Винбергом, обычно трактуется как распределение энергии, аккумулированной в виде первичной продукции, между всеми остальными верхними трофическими уровнями: зоо- и бактериопланктоном, бентосными организмами, рыбами, водными и околоводными животными и человеком. Анализ биотических балансов для разных водоемов позволил выявить ряд общих закономерностей и выводов [Алимов, 1989]. Например:
Мы привели эти цифры, чтобы показать некоторую механистическую условность биотических балансов экосистем, основанных исключительно на энергетике сгорания органического вещества (напомним, что калорийность биомассы равна количеству теплоты, выделившейся при ее сжигании до воды и углекислоты). В результате этого, расходная часть (т.е. вторичная продукция) энергетического баланса водоема оказывается существенно меньше приходной части (первичной продукции) – см. табл. 21 [Алимов, 1989]. При этом не учитывается, что при последовательном переходе на более высокие трофические уровни биохимическая материя тела особей приобретает дополнительную ценность, хотя бы по причине эндотермичности процессов синтеза высокорганизованного белка в процессе роста. Если проиллюстрировать это отвлеченным примером, то вязанка дров и мешок с долларовыми ассигнациями может иметь одинаковую теплотворную способность, но потребительская и энергетическая (в смысле затрат на их изготовление) ценность долларов значительно выше. На наш взгляд, оценивать в балансах продукцию рыб и других консументов исключительно как "топливо" в терминах теплоты сгорания без учета энергии образования, затраченной на синтетические биохимические процессы, представляется не вполне корректным.
В научном сообществе к проблеме биотических балансов водных экосистем имеется далеко не однозначное отношение, иллюстрируемое следующим "декларативным мемом":
Мем № 26
: “Работы Г.Г. Винберга и А.А. Умнова, на которые ссылается автор, были, прежде всего, вызваны требованиями унификации подходов к обработке и представлению многочисленных данных по продукции, обмену, рационам и другим биоэнергетическим параметрам, накопленных в рамках МБП в различных академических и отраслевых институтах. В те годы действительно господствовало мнение, что "крупноблочные" балансовые модели приблизят нас к управлению потоками энергии в естественных и искусственных (в основном водных) экосистемах. Потребовалось гораздо менее 26 лет, чтобы понять, что даже успешное сведение баланса (правда редко реализуемое) констатирует КПД процессов, но не раскрывает их механизмы. Балансовые модели ставили больше вопросов, чем давали ответов и вынудили отказаться от многих упрощенных схем и представлений. Нисколько не умаляя роль этого подхода в активизации научной мысли, следует признать, что он представляет всё же исторический интерес”.[Из рецензии редакции
"Журнала общей биологии", на отклоненную статью В.К. Шитикова, 2002 г.]На наш взгляд, изучение биопродукционных процессов на "микрокинетическом" уровне (особь и группа особей) отнюдь не является "историческим анахронизмом", а представляет собой одну из основных задач гидробиологии. Что касается "крупноблочных" балансовых моделей, то серьезный и беспристрастный анализ опыта их разработки с доказательной оценкой уровня агрегированности (популяция, трофический уровень или водоем в целом), выше которой любые балансы делаются бессмысленными, еще ждет своего читателя. В любом случае, несмотря на декларации рецензентов редакции "Журнала общей биологии", в пользу "макрокинетических" биотических балансов говорят конкретные практические результаты, позволившие, например, А.Ф. Алимову, Н.М. Мингазовой и другим исследователям дать достаточно детальную оценку состояния экосистем уникальных карстовых водоемов Татарстана [Уникальные экосистемы.., 2001]. Караван продолжает идти...
Иллюстративный материал
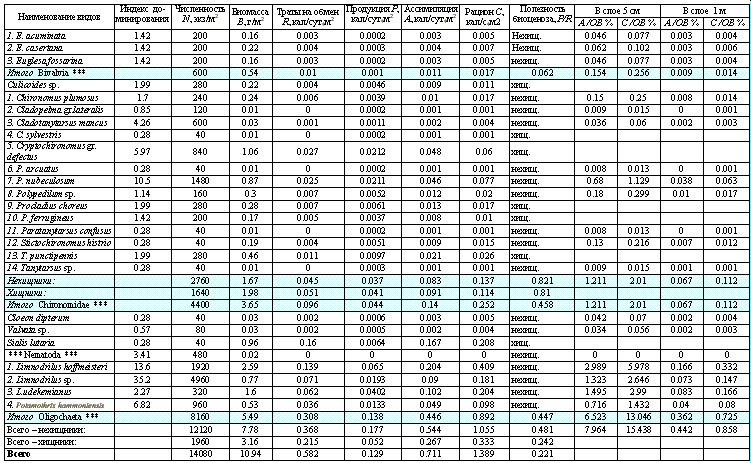
Приведем практический пример расчета составляющих биопродукционного баланса сообщества зообентоса на одной из станций на р. Чапаевка, выполненный по результатам разовых экспедиционных наблюдений и с использованием упрощенной схемы. При отсутствии данных о первичной продукции, популяционной динамики донных организмов и их трофической связи с другими консументами, представленные расчеты являются в значительной мере условными и приближенными. Результаты представим в сводной табл. 4.12 (все энергетические показатели приведены в ккал).
Траты на обмен у каждого i-го вида донных животных рассчитаем по уравнениям потребления кислорода животными в процессе дыхания в зависимости от массы тела по следующей формуле, включающей оксикалорийные и пересчетные коэффициенты:
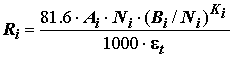
где:
Ni и Bi – численность и биомасса рассчитываемого вида гидробионта; Ai и Ki – коэффициенты степенного уравнения энергетического обмена для данной группы животных по вышеприведенным литературным данным, а также по работам Е.В. Балушкиной [1987], С.М. Голубкова [2000] и др.; e t – температурная поправка на ускорение обменных процессов по уравнению Вант-Гоффа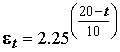 , t – температура воды в придонном слое в момент отбора пробы.
, t – температура воды в придонном слое в момент отбора пробы.
Продукцию популяций отдельных видов животных рассчитаем по известным из литературы величинам удельной продукции за сутки Сw [Заика, 1983]. Величина
Cw равна для олигохет 0.03, личинок хирономид – 0.033, личинок поденок – 0.027, моллюсков сфереид – 0.0044 и др. [Алимов, Финогенова, 1976]. Для дальнейших расчетов используем данные по калорийности (ккал/г) разных групп животных, что позволяет перевести весовые значения составляющих энергобаланса в энергетические: для олигохет – 1.0, хирономид – 0.63, поденок – 0.92, сфериид с раковиной – 0.35, гаммарид – 0.9, вислокрылок – 0.92 [Алимов с соавт., 1977; Каменев, 1993].В каждом донном сообщесте выделим два трофических уровня – нехищных и хищных животных. Ко вторым отнесем личинок вислокрылок, хелеид и некоторых видов хирономид (
Ablabesmyia longistyla, A. monilis, Cryptochironomus gr. defectus, Procladius choreus, Pr. ferrugineus, Tanypus punctipennis).Суммарную продукцию донных сообществ (
Pc) определим в виде суммы продукций нехищных (Pнх) и хищных (Pх) животных за вычетом рациона хищников (Cх): Pс = Pнх + Pх - Cх [Алимов, 1989]. В связи с тем, что основную долю хищников в р. Чапаевка составляют личинки хирономид, которые являются в основном факультативными хищниками, рацион хищных животных для дальнейших расчетов примем равным 30% [Тодераш, 1984]. Тогда уравнение расчета продукции донных сообществ будет иметь следующий вид:Pс = Pнх + Pх – 0.30 × Pх .
Для каждого вида и таксономической группы рассчитаем также отношение продукции донных сообществ к суммарным тратам на обменные процессы Pс
/R, которое рассматривается как коэффициент стабильности или полезного действия донных сообществ на различных участках реки [Алимов, Финогенова, 1976].Биоразнообразие анализируемых донных сообществ оценим по индексу Шеннона, а характер доминирования отдельных видов – в соответствии с индексом по В.Ф. Палию и А. Ковнацкому.
Полученные результаты по составляющим энергетического баланса донных сообществ могут быть использованы в дальнейшем для количественной оценки их участия в процессах самоочищения водоемов. Калорийность (энергосодержание) органического вещества ОВ в воде рассчитывается по величине бихроматной окисляемости БХО (гO
2/м3) для двух вариантов:ОВ
ОВ
max = Ков ОВ× ккал/м2 – в слое 1 м,где Ков
= 0.67× 10.6× 0.5 = 3.551 ккал/г – пересчетный коэффициент теплотворной способности [Алимов с соавт., 1977].Итог самоочищающей способности донных организмов оценим как отношение ассимилированной энергии (А = Р + R) и суточного рациона С нехищных животных зообентоса, к калорийности органического вещества ОВ, распределенного в слое воды 5 см и 1 м над 1 м
2 площади дна. В результате расчетов по данному примеру можно увидеть, что донное сообщество ежесуточно утилизирует свыше 15% чужеродных (ксенобиотических) органических веществ, содержащихся в придонном слое воды, причем около 8% общей массы ОВ расходуется непосредственно на прирост биомассы и обменные процессы. Доля зообентоса в самоочищении реки для слоя воды 1 м существенно меньше, т.к. в средних и верхних слоях водоема ведущая роль в деструкции загрязнений переходит к планктонным организмам и бактериальной микрофлоре, продукция которых нами не учитывалась.Несмотря на известные допущения при расчете биотического баланса, имеющие место в описанном иллюстративном примере, эта методика позволила нам в свое время обработать несколько сотен проб зообентоса и представить достаточно четкую картину антропогенного загрязнения р. Чапаевка, оценить пространственную динамику функционально-продукционных характеристик сообщества донных организмов и долю его участия в самоочищении реки [Экологическое состояние.., 1997].
Таблица 4.12.
Сводная расчетная таблица гидробиологических данных
Станция: Чапаевка / Ст.04л 1км ниже с. Ореховка. Дата: 24 июля 1990 г.
Температура 22.2 градусов C. Бихроматная окисляемость = 38.5. Глубина = 0.9 м.
Калорийность орг. вещества от ОВ(min) = 6.84 до ОВ(max) = 123.04.
Число видов: 26. Индекс разнообразия Шеннона = 3.384.
| Дальше
|
Назад |
Начало |
Конец |
Список
|